
5.1: Transformación en Bacterias
- Page ID
- 57098

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Las bacterias no tienen reproducción sexual en el sentido que hacen los eucariotas. No tienen alternancia de generaciones diploides y haploides, ni gametos, ni meiosis. Sin embargo, la esencia del sexo es la recombinación genética, y las bacterias sí tienen tres mecanismos para lograrlo: transformación, conjugación y transducción.
Transformación
Muchas bacterias pueden adquirir nuevos genes al tomar moléculas de ADN (por ejemplo, un plásmido) de sus alrededores. La capacidad de transformar deliberadamente la bacteria E. coli ha hecho posible la clonación de muchos genes, incluyendo genes humanos, y el desarrollo de la industria biotecnológica. La primera demostración de transformación bacteriana se realizó con Streptococcus pneumoniae y condujo al descubrimiento de que el ADN es la sustancia de los genes. El camino que condujo a este descubrimiento de época comenzó en 1928 con el trabajo de un bacteriólogo inglés, Fred Griffith.


Las células de S. pneumoniae (también conocidas como neumococo) suelen estar rodeadas por una cápsula gomosa hecha de un polisacárido. Cuando se cultiva en la superficie de un medio de cultivo sólido, la cápsula hace que las colonias tengan un aspecto brillante y suave. Estas células se llaman células “S”. Sin embargo, después de un cultivo prolongado en medio artificial, algunas células pierden la capacidad de formar la cápsula, y la superficie de sus colonias está arrugada y rugosa (“R”). Con la pérdida de su cápsula, las bacterias también pierden su virulencia. La inyección de un solo neumococo S en un ratón matará al ratón en aproximadamente 24 horas. Pero una inyección de más de 100 millones (100 x 10 6) células R es totalmente inofensiva.

¿La razón? La cápsula evita que los neumococos sean engullidos y destruidos por células depuradoras, neutrófilos y macrófagos, en el cuerpo. Las formas R están completamente a merced de los fagocitos. Los neumococos también se presentan en más de 90 tipos diferentes: I, II, III y así sucesivamente. Los tipos difieren en la química de su cápsula de polisacárido. A diferencia del desplazamiento ocasional de S -> R, el tipo de organismo es constante. Los ratones inyectados con algunas células S de, digamos, neumococos Tipo II, pronto tendrán sus cuerpos llenos de células descendientes del mismo tipo.
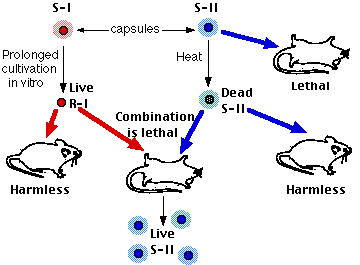
Sin embargo, Griffith encontró que cuando las células R vivas (que deberían haber sido inofensivas) y las células S muertas (que también deberían haber sido inofensivas) se inyectaron juntas, el ratón se enfermó y las células S vivas pudieron recuperarse de su cuerpo. Además, el tipo de células recuperadas del cuerpo del ratón se determinó por el tipo de células S muertas. En el experimento mostrado anteriormente, la inyección de células R-I vivas y células S-II muertas produjo un ratón moribundo con su cuerpo lleno de neumococos S-II vivos. Las células S-II se mantuvieron fieles a su nuevo tipo. Algo en las células S-II muertas había hecho un cambio permanente en el fenotipo de las células R-I. El proceso se denominó transformación.
Oswald Avery y sus colegas en The Rockefeller Institute en la ciudad de Nueva York finalmente demostraron que el “algo” era ADN. Al perseguir el descubrimiento de Griffith, encontraron que podían llevar a cabo el mismo tipo de transformación in vitro usando un extracto de las células bacterianas. Trataron este extracto con:
- una enzima para destruir el polisacárido de la cápsula (S-III en sus experimentos)
- disolventes para eliminar todos los lípidos
- tripsina y quimotripsina para destruir cualquier proteína residual
- RNasa para destruir ARN
Descubrieron que no destruía la capacidad de sus extractos para transformar la bacteria. Sin embargo, tratar los extractos con DNasa para destruir el ADN en ellos sí abolió su actividad transformadora. Entonces el ADN era el único material en las células muertas capaz de transformar células de un tipo a otro. El ADN era la sustancia de los genes.
Aunque la composición química de la cápsula está determinada por genes, la relación es indirecta. El ADN se transcribe en ARN y el ARN se traduce en proteínas. El fenotipo de los neumococos —la composición química de la cápsula de polisacárido— está determinado por las enzimas particulares (proteínas) utilizadas en la síntesis de polisacáridos. Avery y sus colegas Colin MacLeod y Maclyn McCarty publicaron sus hallazgos de época el 1 de febrero de 1944. Desafortunadamente, la importancia de su descubrimiento no fue suficientemente apreciada por los científicos en general y el Comité Nobel en particular, y Avery murió antes de que su trabajo pudiera ser honrado con un Premio Nobel. (Los premios Nobel nunca se entregan póstumamente).
Conjugación
Algunas bacterias, E. coli es un ejemplo, pueden transferir una porción de su cromosoma a un receptor con el que están en contacto directo. A medida que el donante replica su cromosoma, la copia se inyecta en el receptor. En cualquier momento en que el donante y el receptor se separen, la transferencia de genes se detiene. Aquellos genes que lograron el viaje con éxito reemplazan sus equivalentes en el cromosoma del receptor.
La conjugación solo puede ocurrir entre células de tipos de apareamiento opuestos: el donante (o “macho”) porta un factor de fertilidad (F +) y el receptor (“hembra”) no (F −). El factor de fertilidad es un conjunto de genes originalmente adquiridos de un plásmido y ahora integrados en el cromosoma bacteriano. Establece el origen de replicación para el cromosoma. Una porción de F es la “locomotora” que introduce el cromosoma en la célula receptora y el resto es el “furgón”.
En E. coli, aproximadamente un gen atraviesa cada segundo que las células permanecen juntas. (Entonces, toma alrededor de 100 min para que todo el genoma (4377 genes) lo haga. Sin embargo, el proceso se interrumpe fácilmente por lo que es más probable que los genes hospedadores cercanos detrás de los genes F principales (“locomotora”) lo hagan que los que están más atrás. El “furgón” rara vez lo hace así al no recibir un factor F completo, la célula receptora sigue siendo “femenina”.
El ADN que lo atraviesa encuentra la región homóloga en el cromosoma femenino y la reemplaza (por un doble cruce). Al separar deliberadamente las células (en una licuadora de cocina) en diferentes momentos, se puede determinar el orden y el espaciado relativo de los genes. De esta manera, se puede hacer un mapa genético —equivalente a los mapas genéticos de eucariotas—. Sin embargo, aquí los intervalos del mapa son segundos, no centimorgans (cM).
La Figura 5.1.4 muestra el mecanismo de conjugación en células de E. coli donde el “macho” carece de genes funcionales necesarios para sintetizar la vitamina biotina y el aminoácido metionina (Bio −, Met −) por lo que estos deben añadirse a su medio de cultivo. La “hembra” tiene esos genes (Bio +, Met +) pero tiene genes no funcionales (mutantes) que le impiden poder sintetizar los aminoácidos treonina y leucina (Thr −, Leu −), por lo que estos deben ser añadido a su medio de cultivo.
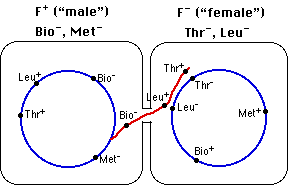
Cuando se cultivan juntas, algunas células femeninas reciben los genes Thr y Leu funcionales del donante masculino. Un doble crossover les permite reemplazar los alelos no funcionales. Ahora las células ahora pueden crecer en un medio “mínimo” que contiene solo glucosa y sales.
Transducción
Los bacteriófagos son virus que infectan a las bacterias. En el proceso de ensamblaje de nuevas partículas de virus, se puede incorporar en ellas algún ADN huésped. La cabeza del virión solo puede contener tanto ADN por lo que estos virus al tiempo que aún pueden infectar nuevas células hospedadoras y pueden ser incapaces de lizarlas. En cambio, el gen bacteriano autoestopista (o genes) pueden insertarse en el ADN del nuevo huésped, reemplazando a los que ya están ahí y dando al huésped un fenotipo alterado. Este fenómeno se llama transducción.
La comprensión de sistemas complejos casi siempre tiene que esperar a desentrañar los detalles de algún sistema más simple. Puede sentir que tratar de averiguar cómo un tipo de neumococo podría convertirse en otro fue una búsqueda sumamente especializada y esotérica. Pero Avery y sus compañeros de trabajo se dieron cuenta del significado más amplio de lo que estaban observando y, en su momento, el resto del mundo científico también lo hizo. Al elegir trabajar con un sistema bien definido: la conversión de formas R de un tipo en formas S de otro tipo, estos investigadores hicieron un descubrimiento que ha revolucionado la biología y la medicina. Intentar comprender el funcionamiento de sistemas complejos entendiendo primero el funcionamiento de sus partes se llama reduccionismo. Algunos científicos (y muchos no científicos) cuestionan el valor del reduccionismo. Favorecen un enfoque holístico enfatizando el funcionamiento del sistema completo.
No obstante, el registro habla por sí mismo. Desde los rascacielos hasta los paseos por la luna, pasando por los chips de computadora hasta los avances de la medicina moderna, el progreso viene de comprender primero las propiedades de las partes que componen el conjunto. El fallecido George Wald, quien ganó el Premio Nobel de Fisiología de 1967 por sus descubrimientos de las bases moleculares de la detección de luz, alguna vez se preocupó de que su trabajo fuera demasiado especializado —estudiando no la visión, ni el ojo, ni toda la retina, ni siquiera sus bastones y conos, sino solo las reacciones químicas de sus rodopsinas. Pero llegó a darse cuenta “es como si se tratara de una ventana muy estrecha a través de la cual a cierta distancia se puede ver sólo una grieta de luz. A medida que uno se acerca, la vista se hace cada vez más amplia, hasta que finalmente a través de esta misma ventana se está mirando al universo. Creo que así es como siempre va en la ciencia, porque la ciencia es toda una. Apenas importa donde uno entre, siempre y cuando uno pueda acercarse más...”.
Importancia de la recombinación genética en bacterias
Transformación, conjugación y transducción fueron descubiertas en el laboratorio. ¿Qué importancia tienen estos mecanismos de recombinación genética en la naturaleza? La finalización de la secuencia de todo el genoma de una variedad de bacterias diferentes (y arqueas) sugiere que los genes en el pasado se han movido de una especie a otra. Este fenómeno se denomina transferencia génica lateral (LGT). La notable propagación de la resistencia a múltiples antibióticos puede haber sido ayudada por la transferencia de genes de resistencia dentro de las poblaciones e incluso entre especies. Muchas bacterias tienen enzimas que les permiten destruir el ADN extraño que ingresa a sus células. Parece poco probable que estos fueran necesarios si eso no ocurriera en la naturaleza. En cualquier caso, estas enzimas de restricción han proporcionado las herramientas de las que dependen los avances de la biología molecular y la industria biotecnológica.