
5.4: Emparejamiento de bases en ADN y ARN
- Page ID
- 57113

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Reglas de Emparejamiento Base
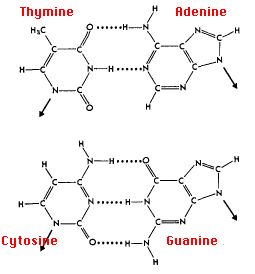
Las reglas del emparejamiento de bases (o emparejamiento de nucleótidos) son:
- A con T: la purina adenina (A) siempre se empareja con la pirimidina timina (T)
- C con G: la pirimidina citosina (C) siempre se empareja con la purina guanina (G)
Esto es consistente con que no hay suficiente espacio (20 Å) para que dos purinas encajen dentro de la hélice y demasiado espacio para que dos pirimidinas se acerquen lo suficiente entre sí para formar enlaces de hidrógeno entre ellas. Pero ¿por qué no A con C y G con T? La respuesta: solo con A & T y con C & G hay oportunidades para establecer enlaces de hidrógeno (mostrados aquí como líneas punteadas) entre ellos (dos entre A & T; tres entre C & G). Estas relaciones a menudo se llaman las reglas del emparejamiento de bases Watson-Crick, que llevan el nombre de los dos científicos que descubrieron su base estructural.
| Organismo | A | T | G | C |
|---|---|---|---|---|
| Humano | 30.9 | 29.4 | 19.9 | 19.8 |
| Pollo | 28.8 | 29.2 | 20.5 | 21.5 |
| Saltamontes | 29.3 | 29.3 | 20.5 | 20.7 |
| Erizo de mar | 32.8 | 32.1 | 17.7 | 17.3 |
| Trigo | 27.3 | 27.1 | 22.7 | 22.8 |
| Levadura | 31.3 | 32.9 | 18.7 | 17.1 |
| E. coli | 24.7 | 23.6 | 26.0 | 25.7 |
Las reglas del emparejamiento de bases nos dicen que si podemos “leer” la secuencia de nucleótidos en una hebra de ADN, podemos deducir inmediatamente la secuencia complementaria en la otra hebra. Las reglas del emparejamiento de bases explican el fenómeno de que sea cual sea la cantidad de adenina (A) en el ADN de un organismo, la cantidad de timina (T) es la misma (llamada regla de Chargaff). De igual manera, sea cual sea la cantidad de guanina (G), la cantidad de citosina (C) es la misma. La relación C+G:A+T varía de organismo a organismo, particularmente entre las bacterias, pero dentro de los límites del error experimental, A=T y C=G.