
15.8A: Neuronas
- Page ID
- 56693

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Las células excitables son aquellas que pueden ser estimuladas para crear una pequeña corriente eléctrica. Las fibras musculares y las células nerviosas (neuronas) son excitables. La foto en color es de una única interneurona en la retina de un conejo. A la célula se le ha inyectado un colorante fluorescente para revelar todas sus ramas. Cada una de las pequeñas perillas en las puntas de las ramas realiza una sinapsis con otra célula en la retina.
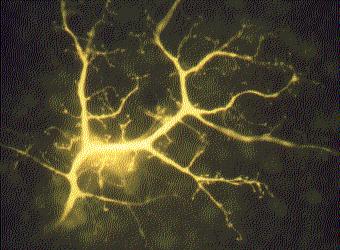
La corriente eléctrica en las neuronas se utiliza para transmitir rápidamente señales a través del animal, mientras que la corriente en los músculos se usa para iniciar la contracción.
El potencial de descanso
Todas las células (no solo las células excitables) tienen un potencial de reposo: una carga eléctrica a través de la membrana plasmática, con el interior de la célula negativa con respecto al exterior. El tamaño del potencial de reposo varía, pero en las células excitables corre alrededor de −70 milivoltios (mv). El potencial de descanso surge de dos actividades:
- La ATPasa sodio/potasio: Esta bomba empuja solo dos iones potasio (K +) dentro de la celda por cada tres iones de sodio (Na +) bombea fuera de la celda por lo que su actividad resulta en una pérdida neta de cargas positivas dentro de la célula.
- Algunos canales de potasio en la membrana plasmática son “permeables” permitiendo una lenta difusión facilitada de K + fuera de la célula (flecha roja).
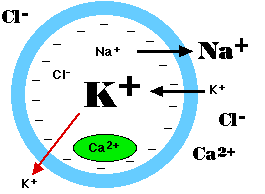
Relaciones Iónicas en la Célula
La ATPasa de sodio/potasio produce
- una concentración de Na + fuera de la célula que es unas 10 veces mayor que la del interior de la célula
- una concentración de K + dentro de la célula unas 20 veces mayor que la que se encuentra fuera de la célula.
Las concentraciones de iones cloruro (Cl −) e iones calcio (Ca 2+) también se mantienen a mayores niveles fuera de la célula EXCEPTO que algunos compartimentos intracelulares encerrados en la membrana también pueden tener altas concentraciones de Ca 2+ ( óvalo verde).
Despolarización
Ciertos estímulos externos reducen la carga a través de la membrana plasmática.
- estímulos mecánicos (por ejemplo, estiramiento, ondas sonoras) activan canales de sodio mecánicamente cerrados
- ciertos neurotransmisores (por ejemplo, acetilcolina) abren los canales de sodio activados por ligando
En cada caso, la difusión facilitada de sodio en la célula reduce el potencial de reposo en ese punto de la célula creando un potencial postsináptico excitatorio o EPSP. Si el potencial se reduce al voltaje umbral (aproximadamente −50 mv en neuronas de mamíferos), se genera un potencial de acción en la célula.
Potenciales de acción
Si la despolarización en un punto de la celda alcanza el voltaje umbral, el voltaje reducido ahora abre cientos de canales de sodio dependientes de voltaje en esa porción de la membrana plasmática. Durante el milisegundo que los canales permanecen abiertos, unos 7000 Na + se precipitan hacia la celda. La despolarización completa repentina de la membrana abre más canales de sodio activados por voltaje en porciones adyacentes de la membrana. De esta manera, una onda de despolarización barre a lo largo de la célula. Este es el potencial de acción. (En las neuronas, el potencial de acción también se llama impulso nervioso).
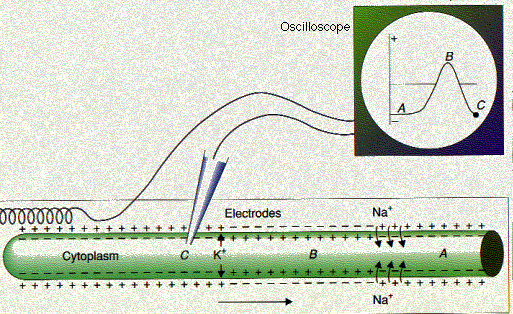
El impulso nervioso: (Figura 15.8.1.3) En la neurona en reposo, el interior de la membrana axónica está cargado negativamente con respecto al exterior (A). A medida que el potencial de acción pasa (B), la polaridad se invierte. Entonces la salida de iones K + restaura rápidamente la polaridad normal (C). En el instante representado en el diagrama, el punto móvil, que ha rastreado estos cambios en el osciloscopio a medida que el impulso barrió más allá del electrodo intracelular, se encuentra en la posición C.
El periodo refractario
Un segundo estímulo aplicado a una neurona (o fibra muscular) menos de 0.001 segundos después del primero no desencadenará otro impulso. La membrana está despolarizada (posición B) y la neurona se encuentra en su periodo refractario. No hasta que se restablezca la polaridad de −70 mv (posición C) la neurona estará lista para volver a disparar. La repolarización se establece primero por la difusión facilitada de iones de potasio fuera de la célula. Sólo cuando la neurona finalmente está descansada son los iones de sodio que entraron en cada impulso se transportan activamente de vuelta fuera de la célula. En algunas neuronas humanas, el periodo refractario dura solo 0.001—0.002 segundos. Esto significa que la neurona puede transmitir 500—1000 impulsos por segundo.
El potencial de acción es todo o ninguno
La fuerza del potencial de acción es una propiedad intrínseca de la célula. Mientras puedan alcanzar el umbral de la célula, los estímulos fuertes no producen potenciales de acción más fuertes que los débiles. Sin embargo, la fuerza del estímulo se codifica en la frecuencia de los potenciales de acción que genera.
Neuronas Mieladas
Los axones de muchas neuronas están encerrados en una vaina grasa llamada vaina de mielina. Es la membrana plasmática muy expandida de una célula accesoria llamada célula de Schwann. Donde la vaina de una célula de Schwann se encuentra con la siguiente, el axón está desprotegido. Los canales de sodio activados por voltaje de las neuronas mielinizadas están confinados a estas manchas (llamadas nódulos de Ranvier).

La irrupción de iones de sodio en un nodo crea la despolarización suficiente para alcanzar el umbral del siguiente. De esta manera, el potencial de acción salta de un nodo al siguiente. Esto da como resultado una propagación del impulso nervioso mucho más rápida de lo que es posible en las neuronas no mielinizadas.
Esclerosis múltiple
Este trastorno autoinmune resulta en la destrucción gradual de las vainas de mielina. A pesar de esto, la transmisión de impulsos nerviosos continúa por un período ya que la célula inserta canales adicionales de sodio activados por voltaje en porciones de la membrana anteriormente protegidas por mielina.
Hiperpolarización
A pesar de su nombre, algunos neurotransmisores inhiben la transmisión de impulsos nerviosos. Lo hacen abriendo canales de cloruro y/o canales de potasio en la membrana plasmática. En cada caso, la apertura de los canales aumenta el potencial de membrana al dejar que los iones cloruro cargados negativamente (Cl −) IN y los iones de potasio cargados positivamente (K +) OUT. Esta hiperpolarización se denomina potencial postsináptico inhibitorio (IPSP) porque contrarresta cualquier señal excitadora que pueda llegar a esa neurona. Aunque el voltaje umbral de la célula no cambia, ahora requiere de un estímulo excitatorio más fuerte para alcanzar el umbral.
Ejemplo: Ácido gamma aminobutírico (GABA). Este neurotransmisor se encuentra en el cerebro e inhibe la transmisión nerviosa por ambos mecanismos:
- la unión a los receptores GABA A abre los canales de cloruro en la neurona
- la unión a los receptores GABA B abre los canales de potasio
Integrando señales
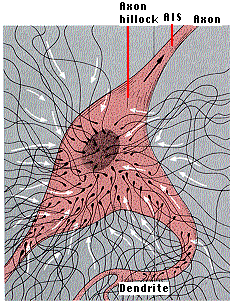
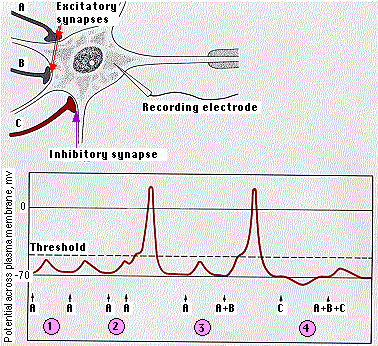
Una sola neurona, especialmente una en el sistema nervioso central (ver foto en color en la parte superior), puede tener miles de otras neuronas sinapsionando en ella. Algunos de estos liberan neurotransmisores activadores (despolarizantes); otros liberan neurotransmisores inhibidores (hiperpolarizantes).
La célula receptora es capaz de integrar estas señales. El diagrama muestra cómo funciona esto en una neurona motora.
- La EPSP creada por una sola sinapsis excitatoria es insuficiente para alcanzar el umbral de la neurona.
- Los EPSP creados en rápida sucesión, sin embargo, se suman (” suma “). Si alcanzan el umbral, se genera un potencial de acción.
- Los EPSP creados por sinapsis excitatorias separadas (A + B) también se pueden sumar para alcanzar el umbral.
- La activación de las sinapsis inhibitorias (C) hace que el potencial de reposo de la neurona sea más negativo. El IPSP resultante también puede evitar que lo que de otro modo habría sido EPSP efectivos desencadene un potencial de acción.
Normalmente, el número de EPSP necesarios para alcanzar el umbral es mayor que el que se muestra aquí.
Se podría esperar que la despolarización en un punto de la membrana plasmática genere un potencial de acción independientemente de las señales inhibitorias en otros lugares. Sin embargo, esto se evita en muchas neuronas por el montículo del axón y el segmento inicial del axón (el AIS). Esta es la región donde el axón emerge del cuerpo celular y se encuentra sin mielinizar. La porción de la membrana plasmática en esta región tiene pocas o ninguna sinapsis propias y un umbral más bajo que en otras partes de la célula.
Las neuronas pueden establecer dichos dominios distintivos en su membrana plasmática anclando (con filamentos de actina) proteínas transmembrana como barreras para bloquear la difusión libre de proteínas de membrana desde el cuerpo celular hasta el axón. El potencial de acción generalmente se genera en el segmento inicial del axón. Al no tener sinapsis excitadoras ni inhibidoras propias, es capaz de evaluar la imagen total de las EPSP e IPSP creadas en las dendritas y el cuerpo celular. Solo si, en un breve intervalo, la suma de las señales despolarizantes menos la suma de las señales hiperpolarizantes excede el umbral del segmento inicial del axón, se generará un potencial de acción.
Este método para que la neurona evalúe una mezcla de señales positivas y negativas ocurre rápidamente. Resulta, sin embargo, que las neuronas también tienen una forma a largo plazo de integrar una mezcla de señales positivas y negativas que convergen en ellas. Esta respuesta a largo plazo implica cambios en la actividad génica que conducen a cambios en el número y la actividad de las muchas sinapsis de la célula.