
15.9C: Visión
- Page ID
- 56996

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
El ojo humano está envuelto en tres capas de tejido:
- el pelaje esclerótico. Esta capa dura crea el “blanco” del ojo excepto en la parte frontal donde forma la córnea transparente. La córneaLa superficie de la córnea se mantiene húmeda y libre de polvo por las secreciones de las glándulas lagrimales.
- admite luz al interior del ojo y
- dobla los rayos de luz para que puedan ser llevados a un foco.
- el abrigo coroideo. Esta capa media está profundamente pigmentada con melanina. Reduce el reflejo de la luz parásita dentro del ojo. El pelaje coroideo forma el iris en la parte frontal del ojo. Esto, también, está pigmentado y es responsable del “color” de los ojos. El tamaño de su abertura, la pupila, es variable y está bajo el control del sistema nervioso autónomo. Con poca luz (o cuando el peligro amenaza), la pupila se abre más y deja entrar más luz en el ojo. En luz brillante la pupila se cierra. Esto no sólo reduce la cantidad de luz que ingresa al ojo sino que también mejora su capacidad de formación de imágenes (al igual que “deteniendo” el diafragma iris de una cámara).
- la retina La retina es la capa interna del ojo. Contiene los receptores de luz, los bastones y conos (y así sirve como la “película” del ojo). La retina también tiene muchas interneuronas que procesan las señales que surgen en los bastones y conos antes de pasarlas de regreso al cerebro. (Nota: los bastones y conos no están en la superficie de la retina sino que se encuentran debajo de la capa de interneuronas.)

Todos los impulsos nerviosos generados en la retina viajan de regreso al cerebro por medio de los axones en el nervio óptico (arriba). En el punto de la retina donde los aproximadamente 1 millón de axones convergen sobre el nervio óptico, no hay bastones ni conos. Este punto, llamado punto ciego, es así insensible a la luz.

Se puede demostrar la presencia del punto ciego. Cúbrete el ojo derecho con la mano y mira fijamente el círculo rojo a medida que te acercas a la pantalla (el cuadrado desaparecerá). O cúbrete el ojo izquierdo y mira fijamente el cuadrado rojo mientras te mueves.
La lente
La lente se encuentra justo detrás del iris. Se mantiene en posición por zónulas que se extienden desde un anillo circundante de músculo. Cuando este músculo ciliar se relaja, su diámetro aumenta, las zónulas se ponen bajo tensión, y el cristalino se aplana y cuando se contrae, su diámetro se reduce, las zónulas se relajan y el cristalino se vuelve más esférico. Estos cambios permiten que el ojo ajuste su enfoque entre objetos lejanos y objetos cercanos.

Hipermetropía. Si el globo ocular es demasiado corto o el cristalino demasiado plano o inflexible, los rayos de luz que ingresan al ojo —particularmente los de objetos cercanos— no serán enfocados en el momento en que golpeen la retina. Las gafas con lentes convexas pueden corregir el problema. La hipermetropía se llama hipermetropía.
Miopía. Si el globo ocular es demasiado largo o el cristalino demasiado esférico, la imagen de objetos distantes se enfoca frente a la retina y vuelve a estar fuera de foco antes de que la luz incida en la retina. Los objetos cercanos se pueden ver más fácilmente. Las gafas con lentes cóncavas corrigen este problema al divergir los rayos de luz antes de que entren al ojo. La miopía se llama miopía.
Cataratas Uno o ambos lentes suelen volverse turbios a medida que uno envejece. Cuando una catarata interfiere seriamente con la visión, la lente turbia se retira fácilmente y se sustituye por una de plástico. Todo el proceso se puede hacer en pocos minutos como paciente ambulatorio bajo anestesia local.
El iris y el cristalino dividen el ojo en dos cámaras principales:
- la cámara frontal se llena con un líquido acuoso, el humor acuoso
- la cámara trasera está llena de un material gelatinoso, el cuerpo vítreo
La Retina
Cuatro tipos de receptores sensibles a la luz se encuentran en la retina:
- varillas
- tres tipos de conos, cada uno “sintonizado” para responder mejor a la luz de una porción del espectro de luz visible
- conos que absorben luz de longitud de onda larga (rojo)
- conos que absorben luz de longitud de onda media (verde)
- conos que absorben luz de longitud de onda corta (azul)
Esta micrografía electrónica de barrido (cortesía de Scott Mittman y David R. Copenhagen) muestra bastones y conos en la retina de la salamandra tigre. Cada tipo de receptor tiene su propio pigmento especial para absorber la luz. Cada uno consiste en una proteína transmembrana llamada opsina acoplada al grupo protésico retiniano. La retina es un derivado de la vitamina A (lo que explica por qué la ceguera nocturna es un signo de deficiencia de vitamina A) y es utilizada por los cuatro tipos de receptores.

La secuencia de aminoácidos de cada uno de los cuatro tipos de opsina es similar, pero las diferencias explican sus diferencias en el espectro de absorción. La retina también contiene una compleja matriz de interneuronas:
- células bipolares y células ganglionares que juntas forman un camino desde los bastones y conos hasta el cerebro
- una compleja matriz de otras interneuronas que forman sinapsis con las células bipolares y ganglionares y modifican su actividad.
Las células ganglionares siempre están activas. Incluso en la oscuridad generan trenes de potenciales de acción y los conducen de regreso al cerebro a lo largo del nervio óptico. La visión se basa en la modulación de estos impulsos nerviosos. No existe la relación directa entre el estímulo visual y un potencial de acción que se encuentra en los sentidos del oído, el gusto y el olfato. De hecho, los potenciales de acción ni siquiera se generan en las barras y conos.
Visión de Rod
La rodopsina es el pigmento que absorbe la luz de las varillas. Este receptor acoplado a proteína G (GPCR) se incorpora en las membranas de discos que están cuidadosamente apilados (unos 1000 o más de ellos) en la porción externa de la varilla. Esta disposición recuerda a la organización de los tilacoides, otro dispositivo que absorbe la luz.

Fig.15.9.3.5 Células de varilla de murciélago canguro
La micrografía electrónica (cortesía de Keith Porter) muestra las células bastoncillas de la rata canguro. Los segmentos externos de las varillas contienen las pilas ordenadas de membranas que incorporan rodopsina. La porción interna contiene muchas mitocondrias. Las dos partes de la varilla están conectadas por un tallo (flecha) que tiene la misma estructura que un cilio primario. Aunque los discos se forman inicialmente a partir de la membrana plasmática, se separan de ella. Así, las señales generadas en los discos deben ser transmitidas por un mediador químico (un “segundo mensajero” llamado GMP cíclico (cGMP) para alterar el potencial de la membrana plasmática de la varilla. La rodopsina consiste en una opsina de 348 aminoácidos acoplada a retinal. Al igual que todos los receptores acoplados a proteínas G, la opsina tiene 7 segmentos de hélice alfa que pasan de un lado a otro a través de la bicapa lipídica de la membrana del disco.
Retina
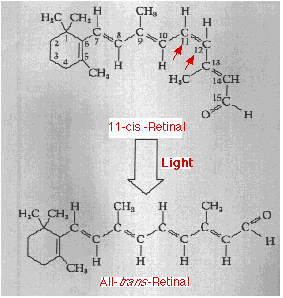
La retina consiste en un sistema de enlaces simples y dobles alternantes. En la oscuridad, los átomos de hidrógeno unidos a los átomos de carbono #11 y #12 de la retina (flechas rojas) apuntan en la misma dirección produciendo una torcedura en la molécula. Esta configuración se designa cis. Cuando la luz es absorbida por la retina, la molécula se endereza formando el isómero all- trans.
![]()
Este cambio físico en la retina desencadena la siguiente cadena de eventos que culminan en un cambio en el patrón de impulsos enviados de vuelta a lo largo del nervio óptico.
- La formación de todo- trans retinal activa su opsina.
- La rodopsina activada, a su vez, activa muchas moléculas de una proteína G llamada transducin.
- La transducin activa una enzima que descompone el GMP cíclico.
- La caída en cGMP cierra los canales de Na + en la membrana plasmática de la varilla. Debido a que estos iones positivos ya no pueden entrar, el interior de la célula se vuelve más negativo (hiperpolarizado) aumentando su potencial de membrana de aproximadamente −30 a unos −70 mV.
- Esto ralentiza la liberación del neurotransmisor glutamato en las sinapsis entre la varilla y las interneuronas (por ejemplo, células bipolares).
- Esta reducción en la liberación de glutamato activa algunas vías interneuronales, inhibe otras.
- La interacción de interneuronas excitadas e inhibidas modula el disparo espontáneo de las células ganglionares a las que están conectadas y da lugar a la capacidad de la retina para discriminar formas.
Entonces la retina no es simplemente una lámina de fotocélulas, sino un minúsculo centro cerebral que lleva a cabo un complejo procesamiento de información antes de enviar señales de vuelta a lo largo del nervio óptico. De hecho, la retina realmente es parte del cerebro y crece de ella durante el desarrollo embrionario.
La visión de varilla es aguda pero gruesa
Las varillas no proporcionan una imagen nítida por varias razones.
- Las varillas adyacentes están conectadas por uniones de hueco y, por lo tanto, comparten sus cambios en el potencial de membrana.
- Varias varillas cercanas suelen compartir un solo circuito con una célula ganglionar.
- Una sola varilla puede enviar señales a varias células ganglionares diferentes.
Entonces, si solo se estimula una sola varilla, el cerebro no tiene forma de determinar exactamente en qué parte de la retina se encontraba. Sin embargo, las varillas son extremadamente sensibles a la luz. Un solo fotón (la unidad mínima de luz) absorbido por un pequeño grupo de varillas adyacentes es suficiente para enviar una señal al cerebro. Entonces, aunque las varillas nos proporcionan una imagen relativamente granulada, incolora, nos permiten detectar una luz que es más de mil millones de veces más tenue que la que vemos en un día soleado y brillante.
Visión de cono
Aunque los conos solo operan con luz relativamente brillante, nos proporcionan nuestras imágenes más nítidas y nos permiten ver colores. La mayoría de los 3 millones de conos en cada retina están confinados a una pequeña región justo enfrente del cristalino llamada fóvea. Por lo que nuestras imágenes más nítidas y coloridas se limitan a un área de visión pequeña. Debido a que rápidamente podemos dirigir nuestros ojos hacia cualquier cosa a la vista que nos interese, tendemos a no ser conscientes de lo pobre que es nuestra visión periférica.
Los tres tipos de conos nos proporcionan la base de la visión del color. Los conos están “sintonizados” a diferentes porciones del espectro visible.
- conos absorbentes rojos; aquellos que absorben mejor a las longitudes de onda relativamente largas que alcanzan un pico a 565 nm
- conos absorbentes verdes con un pico de absorción a 535 nm
- conos absorbentes azules con un pico de absorción a 440 nm.
La retina es el grupo protésico para cada pigmento. Las diferencias en la secuencia de aminoácidos de sus opsinas explican las diferencias en la absorción. La respuesta de los conos no es de todo o ninguno. La luz de una longitud de onda dada (color), digamos 500 nm (verde), estimula los tres tipos de conos, pero los conos que absorben el verde serán estimulados con mayor fuerza. Al igual que las varillas, la absorción de luz no activa potenciales de acción sino que modula el potencial de membrana de los conos.
Ceguera al color
El término daltonismo es algo así como un nombre erróneo. Muy pocas (~1 de cada 10 5) personas no pueden distinguir los colores en absoluto. La mayoría de las personas “daltónicas” en realidad tienen visión de color anormal, como confundir el rojo y el verde de los semáforos. Tan alto como 8% de los machos en algunas poblaciones tienen un defecto heredado en su capacidad de discriminar rojos y verdes. Estos defectos se encuentran casi exclusivamente en los machos debido a que los genes que codifican las opsinas que absorben el rojo y el verde se encuentran en el cromosoma X.

El cromosoma X normalmente porta un grupo de 2 a 9 genes de opsina. La base mínima para la visión rojo-verde normal es un gen cuya opsina absorbe eficientemente en el rojo y otro que absorbe bien en el verde (cromosoma 1 en la figura). Múltiples copias de estos genes también están bien (2 y 3). Los machos con falta de “gen verde” o “gen rojo” son severamente daltónicos (4 y 5). Sin embargo, si todos los genes rojos portan mutaciones (esto rara vez parece ser el caso de los genes verdes —nadie sabe por qué), entonces pueden tener daltonismo rojo-verde que varía de leve a grave dependiendo de las mutaciones particulares involucradas (6). La regla parece ser que cuanto más las mutaciones desplazan el pigmento hacia el verde, más grave es el defecto. Sin embargo, un gran número de mutaciones no siempre producen defectos graves. Múltiples mutaciones en un solo gen pueden compensarse entre sí produciendo solo defectos leves. Y mientras esté presente una copia normal de cada gen, la presencia de genes mutados adicionales rara vez produce un problema grave (7).
¿Por qué algunos machos tienen hasta 9 copias de genes que codifican las opsinas rojas y verdes, cuando dos son suficientes? Las secuencias de los genes rojo y verde son las mismas al 98% de sus nucleótidos. Este alto grado de similitud crea el riesgo de desapareamientos en la sinapsis durante la meiosis con cruce desigual.
Visión azul
La sensibilidad anormal al azul ocasionalmente ocurre en humanos pero es mucho más rara que las anomalías en la visión rojo-verde. El gen para la opsina de cono azul se localiza en el cromosoma 7. Así, este rasgo muestra un patrón autosómico de herencia que se encuentra en las hembras con tanta frecuencia como en los machos.