
15.11.8: Ritmos circadianos en Drosophila y Mamíferos
- Page ID
- 56901

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Todos los eucariotas y algunos microbios (por ejemplo, cianobacterias) muestran cambios en la actividad génica, bioquímica, fisiología y comportamiento que se desvanecen y disminuyen a través del ciclo de días y noches.
Ejemplos:
- El nivel de la hormona melatonina que sube en tu cuerpo durante la noche y baja durante el día.
- Las moscas de la fruta (Drosophila melanogaster) eclosionan en mayor número justo al amanecer.
Incluso cuando el organismo se coloca en condiciones constantes (e.g., oscuridad continua), estos ritmos persisten. Sin embargo, sin señales ambientales, tienden a ser algo más largas o algo más cortas que las 24 horas dando lugar al nombre de ritmos circadianos (L. circa = aproximadamente; dies = día).
La genética y la biología molecular de los ritmos circadianos se han estudiado en varios organismos modelo, entre ellos
- algunos eucariotas unicelulares
- hongos
- plantas (Arabidopsis)
- invertebrados (Drosophila)
- mamíferos (ratones, ratas y humanos)
Lo que ha surgido son algunas similitudes notables en los mecanismos a través de estos diversos grupos. Echemos un vistazo detallado al mecanismo en Drosophila.
El reloj circadiano en Drosophila
Una serie de genes en Drosophila se encienden cuando el animal se expone a la luz:
- genes efectores cuyos productos median las respuestas del animal (por ejemplo, eclosión o muda)
- genes de reloj cuyos productos regulan el reloj circadiano. Dos miembros clave de este grupo son:
- periodo (por)
- atemporal (tim)

La activación de todos estos genes requiere que sus promotores estén unidos por los factores de transcripción de proteínas
- CLOCK codificado por el gen clock (clk)
- CICLO codificado por el ciclo génico (cyc)
(Los nombres de las proteínas se designarán con letras romanas en mayúscula; los genes que las codifican se indican en minúscula cursiva).
El Mecanismo
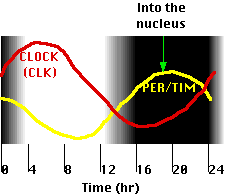
Fig.15.11.8.2 Mecanismo
- Las proteínas PER y TIM (sintetizadas en ribosomas en el citoplasma) forman dímeros.
- Cuando la concentración de estos se eleva lo suficiente (temprano en la noche), se disocian y son transportados al núcleo.
- Aquí PER
- se une a los factores de transcripción CLK/CYC, eliminándolos de los promotores de los genes que activan, cerrando así la transcripción. Debido a que estos genes incluyen per y tim, el resultado es un bucle de retroalimentación negativa; es decir, el producto del per gen inhibe su propia síntesis (así como la de tim). Así como el calor de un horno gira a través del termostato, su propia producción se apaga, así un nivel creciente de dímeros PER/TIM apaga aún más la síntesis de los mismos. A medida que el nivel desciende, esta inhibición se levanta y la actividad PER/TIM comienza de nuevo.
- activa la expresión génica del reloj.
- El tiempo requerido para los diferentes efectos da como resultado que los niveles de PER/TIM y CLOCK oscilen en fases opuestas con un ritmo circadiano (~24 hr) (figura).
Ajuste del reloj
Incluso sin ninguna señal externa (por ejemplo, alternando luz y oscuridad), los ciclos persisten aunque tienden a alejarse del tiempo ambiental.
- En condiciones naturales, los relojes son precisos.
- Esto se debe a que están “fijadas” (sincronizadas) por señales ambientales, de las cuales la luz es una de las más importantes.
En Drosophila, funciona así.
- La luz (azul) es absorbida por la proteína criptocromo (CRY).
- Esto provoca un cambio alostérico en su conformación que le permite unirse a TIM y PER.
- Esto hace que TIM y PER se descompongan (en proteasomas) terminando su inhibición de la transcripción génica.
- Si esto sucede cuando los niveles de PER/TIM están subiendo (tarde en el “día”), hace retroceder el reloj.
- Si sucede cuando los niveles de PER/TIM están disminuyendo (tarde en la “noche”), pone el reloj adelante.
El reloj circadiano en mamíferos
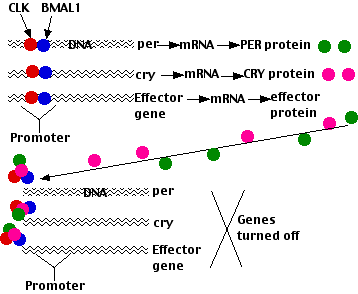
El reloj circadiano en mamíferos se asemeja al de Drosophila de varias maneras con muchos de los genes participantes siendo homólogos. Sin embargo, existen algunas diferencias:
- Los factores de transcripción que encienden los promotores inducidos por la luz son dímeros de la proteína CLOCK y una proteína denominada BMAL1. Estos dímeros se encienden
- tres genes Per
- dos genes Cry, los genes que codifican criptocromo
- cientos de genes efectores cuyos productos controlan una amplia variedad de funciones metabólicas (por ejemplo, respiración celular, glucólisis, gluconeogénesis, metabolismo lipídico)
- Los ARNm PER y CRY se exportan al citoplasma donde se traducen.
- Las proteínas PER y CRY luego ingresan al núcleo donde inhiben CLOCK-BMAL1 desactivando así la transcripción de Per y Cry, y luego se degradan en proteasomas.
A su debido tiempo estas acciones permiten a CLOCK y BMAL1 estimular una vez más la transcripción de Per y Cry. Así, este bucle de retroalimentación negativa hace que los niveles de BMAL1 y PER/CRY oscilen en fases opuestas (como hacen CLOCK y PER/TIM en Drosophila).
Muchos tejidos en mamíferos, por ejemplo, hígado, músculo esquelético y las células beta del páncreas tienen relojes endógenos. Pero todos estos están bajo el control de un “reloj maestro”, el núcleo supraquiasmático (SCN) -cúmulos de neuronas en el hipotálamo.
Los niveles sanguíneos de muchas hormonas tienen fuertes ritmos circadianos. Ejemplos:
- hormonas sintetizadas en el hipotálamo, por ejemplo vasopresina
- cuya secreción es controlada por el hipotálamo como la hormona del crecimiento y el cortisol
- insulina
Ajuste del reloj
Por la luz
Los ratones que son totalmente ciegos (que carecen tanto de bastones como de conos) no tienen problemas para mantener su reloj circadiano a tiempo.
Ellos son capaces de hacer esto porque
- Alrededor del 1— 2% de las células ganglionares en su retina - en lugar de depender de las señales que llegan de bastones y/o conos detectan la luz directamente.
- Estas células ganglionares tienen una extensa red de dendritas que contienen el pigmento melanopsina.
- Cuando se exponen a la luz (la luz difusa es fina), estas células ganglionares se despolarizan y envían sus señales de regreso al núcleo supraquiasmático (SCN).
Por comida
En ratones, el reloj SCN, establecido por ciclos de luz/oscuridad, es el reloj maestro siempre y cuando la comida esté disponible todo el tiempo (la situación normal en el laboratorio). No obstante, si se ofrece comida solo por un periodo de 4 horas en el que los ratones normalmente estarían dormidos, cambian varias actividades circadianas para que, por ejemplo, una vez al día comiencen a correr justo antes de esperar que se les dé comida. Este ritmo continúa aunque los ratones se mantengan en constante oscuridad.
El mecanismo del reloj es el mismo que el reloj impulsado por luz/oscuridad en el SCN, pero la maquinaria que fija el reloj por comida se localiza en una parte diferente del hipotálamo, los núcleos hipotalámicos dorsomediales (DMH).
Los ratones con ambas copias del gen Bmal1 noqueadas, son incapaces de establecer ritmos circadianos ni a la luz ni a los alimentos. Sin embargo, las inyecciones de un vector viral adenoasociado (AAV) que contiene el gen Bmal1
- en el SCN restaura el reloj de luz pero no el reloj de alimentación
- en el DMH restaura la comida pero no el reloj de ajuste de luz.
Trastornos del sueño
A diferencia de los ratones, las personas que son totalmente ciegas no pueden poner el reloj en su SCN. Como resultado, su ritmo circadiano deriva fuera de fase con el ciclo real del día y la noche. A estas personas a menudo les molesta sentirse somnolientas durante el día y bien despiertas cuando quieren estar dormidas por la noche. Un reporte en la edición del 12 de octubre de 2000 del New England Journal of Medicine habla de un grupo de personas ciegas que pudieron ajustar sus relojes con la ayuda de una dosis (10 mg) de melatonina a la hora de acostarse. Sin embargo, este tratamiento funcionó solo cuando el ritmo circadiano del sujeto se había desviado de manera que el aumento normal de melatonina de la glándula pineal se estaba produciendo a primera hora de la noche; es decir, la dosis de melatonina tuvo que darse cuando podría potenciar el nivel endógeno de la hormona.
Algunas personas sufren de un trastorno llamado síndrome familiar avanzado de la fase del sueño (FASPS). Como su nombre indica, se hereda (“familiar”) y sus relojes circadianos corren rápido (“avanzado”). Los afligidos tienden a despertarse varias (hasta cuatro) horas antes de lo normal.
Una de las causas del trastorno resulta ser una mutación puntual en el gen PER2 humano. Exactamente cómo esta mutación provoca el acortamiento del ciclo circadiano aún está bajo investigación.
Fotoperiodismo
Muchas plantas y animales no sólo participan en un ciclo de actividades diarias (apertura de flores, vigilia, alimentación, etc.) sino también en actividades estacionales.
- En las plantas, cosas como la producción de flores y hacer que los cogollos estén latentes en preparación para el invierno.
- En los animales, cosas como prepararse para migrar y entrar y salir de la hibernación.
La pista más confiable para el cambio de estación es la duración del día (¡la temperatura es mucho menos confiable!). Cuanto más lejos viva una planta o animal al norte o al sur del ecuador, más pronunciada será la relación cambiante de las horas diurnas a las horas nocturnas con los cambios de estación.
No es de extrañar entonces que tanto las plantas como los animales dependan principalmente del fotoperiodo para prepararse para los cambios en las actividades estacionales. ¿Y qué mejor manera de medir la duración relativa del día y de la noche que alistando la maquinaria por la que se arrastran los ritmos circadianos?
En cuanto a los animales, un trabajo reciente con Drosophila sugiere que este animal utiliza dos relojes circadianos para monitorear la longitud cambiante del día y la noche.
- Un reloj “vespertino” que toma el relevo en los largos días del verano.
- Un reloj “matutino” que es inhibido por la luz pero que se hace cargo cuando las noches son cada vez más largas;
La maquinaria molecular (Cry, Tim, Per, etc.) para cada reloj está confinada a separar neuronas en dos partes diferentes del cerebro.
En estos experimentos, Drosophila está utilizando los dos relojes para adaptar los ciclos de actividad diarios —no estacionales— a los ciclos de actividad cambiantes de las estaciones. Pero esta maquinaria para medir el fotoperiodo podría permitirles prepararse para los cambios estacionales en la actividad, por ejemplo, dejar de formar huevos al final del verano. Sin embargo, otros estudios que examinan tales cambios estacionales en Drosophila encuentran que la respuesta fotoperiódica es independiente de las respuestas circadianas. Por lo que debemos esperar más experimentos para resolver la cuestión.