
23.3: Grupos de Protistas
- Page ID
- 59898

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Habilidades para Desarrollar
- Describir organismos protistas representativos de cada uno de los seis supergrupos de eucariotas actualmente reconocidos
- Identificar las relaciones evolutivas de plantas, animales y hongos dentro de los seis supergrupos de eucariotas actualmente reconocidos
En el lapso de varias décadas, el Reino Protista ha sido desmontado porque los análisis de secuencias han revelado nuevas relaciones genéticas (y por lo tanto evolutivas) entre estos eucariotas. Además, los protistas que exhiben características morfológicas similares pueden haber desarrollado estructuras análogas debido a presiones selectivas similares, en lugar de por la ascendencia común reciente. Este fenómeno, llamado evolución convergente, es una de las razones por las que la clasificación protista es tan desafiante. El esquema de clasificación emergente agrupa todo el dominio Eukaryota en seis “supergrupos” que contienen a todos los protistas, así como animales, plantas y hongos que evolucionaron a partir de un ancestro común (Figura\(\PageIndex{1}\)). Se cree que los supergrupos son monofiléticos, lo que significa que todos los organismos dentro de cada supergrupo se cree que han evolucionado a partir de un solo ancestro común y, por lo tanto, todos los miembros están más estrechamente relacionados entre sí que con organismos fuera de ese grupo. Aún faltan pruebas para la monofilia de algunos grupos.
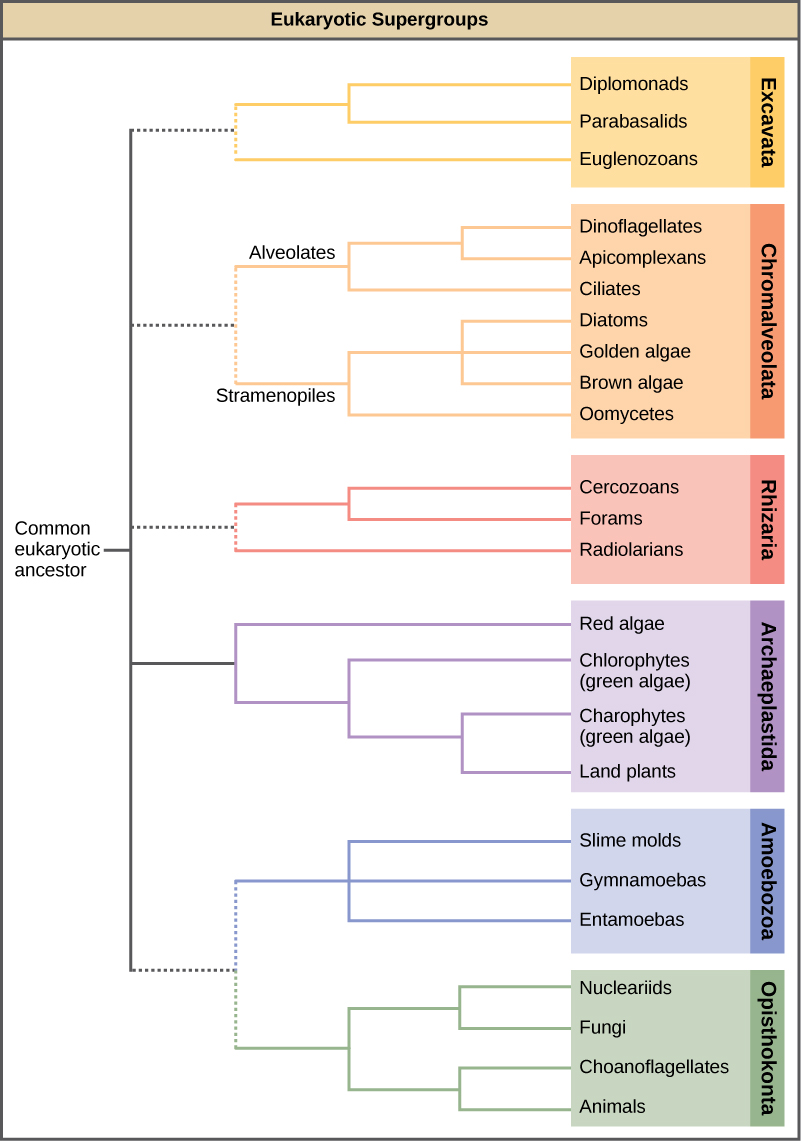
La clasificación de los eucariotas todavía está en cambio, y los seis supergrupos pueden ser modificados o reemplazados por una jerarquía más apropiada a medida que se acumulan datos genéticos, morfológicos y ecológicos. Tenga en cuenta que el esquema de clasificación que aquí se presenta es solo una de varias hipótesis, y las verdaderas relaciones evolutivas aún están por determinar. Al aprender sobre los protistas, es útil enfocarse menos en la nomenclatura y más en los puntos en común y diferencias que definen a los propios grupos.
Excavata
Muchas de las especies protistas clasificadas en el supergrupo Excavata son organismos asimétricos, unicelulares con un surco de alimentación “excavado” de un lado. Este supergrupo incluye depredadores heterótrofos, especies fotosintéticas y parásitos. Sus subgrupos son los diplomonadas, parabasálidos y euglenozoos.
Diplomonadas
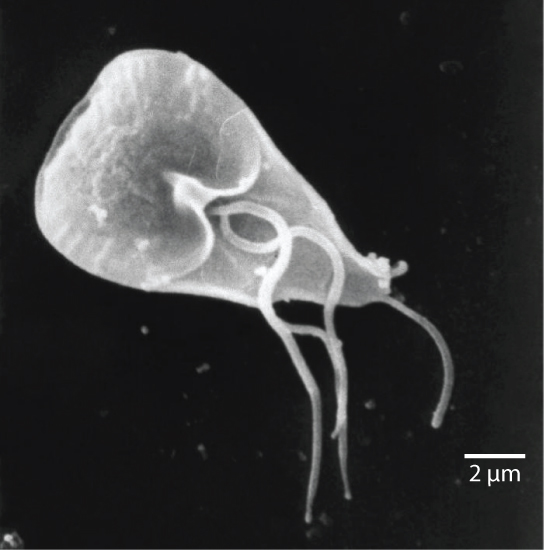
Entre las Excavata se encuentran las diplomonadas, que incluyen al parásito intestinal, Giardia lamblia (Figura\(\PageIndex{2}\)). Hasta hace poco, se creía que estos protistas carecían de mitocondrias. Los orgánulos remanentes mitocondriales, llamados mitosomas, se han identificado desde entonces en diplomonadas, pero estos mitosomas son esencialmente infuncionales. Las diplomonadas existen en ambientes anaeróbicos y utilizan vías alternativas, como la glucólisis, para generar energía. Cada célula diplomonada tiene dos núcleos idénticos y utiliza varios flagelos para la locomoción.
Parabasálidos
Un segundo subgrupo de Excavata, los parabasálidos, también exhibe mitocondrias semifuncionales. En los parabasálidos, estas estructuras funcionan anaeróbicamente y se llaman hidrogenosomas porque producen gas hidrógeno como subproducto. Los parabasálidos se mueven con flagelos y ondulaciones de membrana. Trichomonas vaginalis, parabasalid que causa una enfermedad de transmisión sexual en humanos, emplea estos mecanismos para transitar a través de los tractos urogenitales masculinos y femeninos. T. la vaginalis causa trichamoniasis, la cual aparece en aproximadamente 180 millones de casos en todo el mundo cada año. Mientras que los hombres rara vez presentan síntomas durante una infección con este protista, las mujeres infectadas pueden volverse más susceptibles a la infección secundaria por el virus de la inmunodeficiencia humana (VIH) y pueden tener más probabilidades de desarrollar cáncer de cuello uterino. Mujeres embarazadas infectadas con T. vaginalis tienen un mayor riesgo de complicaciones graves, como el parto prematuro.
Euglenozoos
Los euglenozoos incluyen parásitos, heterótrofos, autótrofos y mixótrofos, que varían en tamaño de 10 a 500 µm. Los euglenoides se mueven a través de sus hábitats acuáticos utilizando dos flagelos largos que los guían hacia fuentes de luz detectadas por un órgano ocular primitivo llamado mancha ocular. El género familiar, Euglena, abarca algunas especies mixotróficas que muestran una capacidad fotosintética solo cuando la luz está presente. En la oscuridad, los cloroplastos de Euglena se encogen y dejan de funcionar temporalmente, y las células en su lugar absorben nutrientes orgánicos de su entorno.
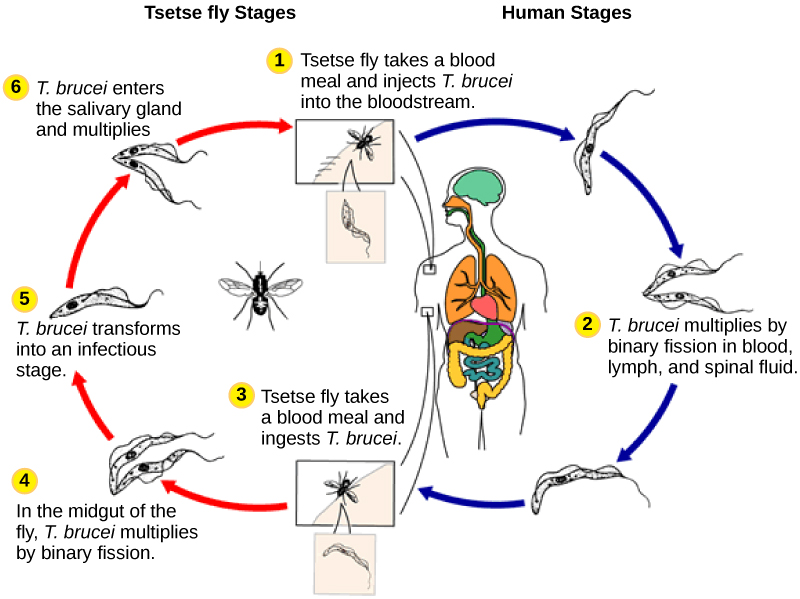
El parásito humano, Trypanosoma brucei, pertenece a un subgrupo diferente de Euglenozoa, los cinetoplastidios. El subgrupo de cinetoplastos lleva el nombre del cinetoplasto, una masa de ADN transportada dentro de la única mitocondria sobredimensionada que posee cada una de estas células. Este subgrupo incluye varios parásitos, llamados colectivamente tripanosomas, que causan enfermedades humanas devastadoras e infectan a una especie de insecto durante una parte de su ciclo de vida. T. brucei se desarrolla en el intestino de la mosca tsé después de que la mosca muerde a un ser humano infectado u otro huésped mamífero. Luego, el parásito viaja a las glándulas salivales del insecto para ser transmitido a otro ser humano u otro mamífero cuando la mosca tsetsé infectada consume otra harina de sangre. T. brucei es común en África central y es el agente causante de la enfermedad africana del sueño, una enfermedad asociada con fatiga crónica severa, coma, y puede ser fatal si no se trata.
Trypanosoma brucei
Mira este video para ver a T. brucei nadando. https://youtu.be/EnsydwITLYk
Chromalveolata
La evidencia actual sugiere que las especies clasificadas como cromalveolatos se derivan de un ancestro común que engulló una célula de algas rojas fotosintéticas, que a su vez ya había evolucionado cloroplastos a partir de una relación endosimbiótica con un procariota fotosintético. Por lo tanto, se cree que el antepasado de los cromalveolatos resultó de un evento endosimbiótico secundario. Sin embargo, algunos cromalveolatos parecen haber perdido orgánulos plastidios derivados de algas rojas o carecen de genes plastidios por completo. Por lo tanto, este supergrupo debe ser considerado un grupo de trabajo basado en hipótesis que está sujeto a cambios. Los cromalveolatos incluyen organismos fotosintéticos muy importantes, como diatomeas, algas pardas y agentes patógenos significativos en animales y plantas. Los cromalveolados se pueden subdividir en alveolados y estramenopiles.
Alveolados: Dinoflagelados, Apicomplexianos y Ciliados
Un gran cuerpo de datos sostiene que los alveolados se derivan de un ancestro común compartido. Los alveolados reciben el nombre de la presencia de un alvéolo, o saco cerrado en la membrana, debajo de la membrana celular. Se desconoce la función exacta del alvéolo, pero puede estar involucrado en la osmorregulación. Los alveolados se categorizan en algunos de los protistas más conocidos: los dinoflagelados, los apicomplexanos y los ciliados.
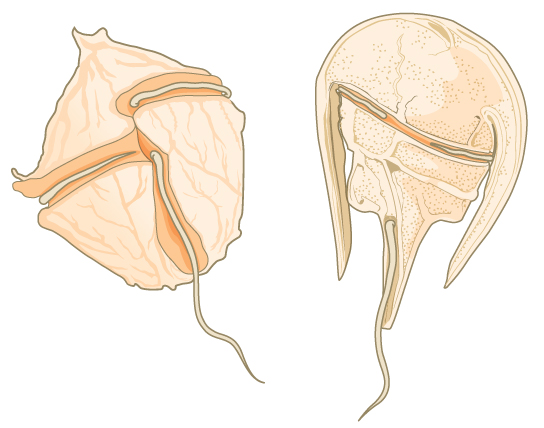
Los dinoflagelados presentan una amplia diversidad morfológica y pueden ser fotosintéticos, heterótrofos o mixotróficos. Muchos dinoflagelados están envueltos en placas entrelazadas de celulosa. Dos flagelos perpendiculares encajan en las ranuras entre las placas de celulosa, con un flagelo que se extiende longitudinalmente y un segundo rodeando el dinoflagelado (Figura\(\PageIndex{4}\)). Juntos, los flagelos contribuyen al característico movimiento de giro de los dinoflagelados. Estos protistas existen en hábitats marinos y de agua dulce, y son un componente del plancton, los organismos típicamente microscópicos que se desplazan a través del agua y sirven como fuente de alimento crucial para los organismos acuáticos más grandes.
Algunos dinoflagelados generan luz, llamada bioluminiscencia, cuando son agitados o estresados. Un gran número de dinoflagelados marinos (miles de millones o billones de células por ola) pueden emitir luz y hacer que toda una ola rompiente brille o adquiera un color azul brillante (Figura\(\PageIndex{5}\)). Para aproximadamente 20 especies de dinoflagelados marinos, las explosiones poblacionales (también llamadas floraciones) durante los meses de verano pueden teñir el océano con un color rojo fangoso. Este fenómeno se llama marea roja, y resulta de los abundantes pigmentos rojos presentes en los plastidios dinoflagelados. En grandes cantidades, estas especies de dinoflagelados secretan una toxina asfixiante que puede matar peces, aves y mamíferos marinos. Las mareas rojas pueden ser enormemente perjudiciales para las pesquerías comerciales, y los humanos que consumen estos protistas pueden llegar a ser envenenados.

Los protistas apicomplejos se llaman así porque sus microtúbulos, fibrina y vacuolas se distribuyen asimétricamente en un extremo de la célula en una estructura llamada complejo apical (Figura\(\PageIndex{6}\)). El complejo apical está especializado para la entrada e infección de células hospedadoras. En efecto, todos los apicomplexanos son parasitarios. Este grupo incluye al género Plasmodium, el cual causa malaria en humanos. Los ciclos de vida apicomplejos son complejos, involucrando múltiples huéspedes y etapas de reproducción sexual y asexual.
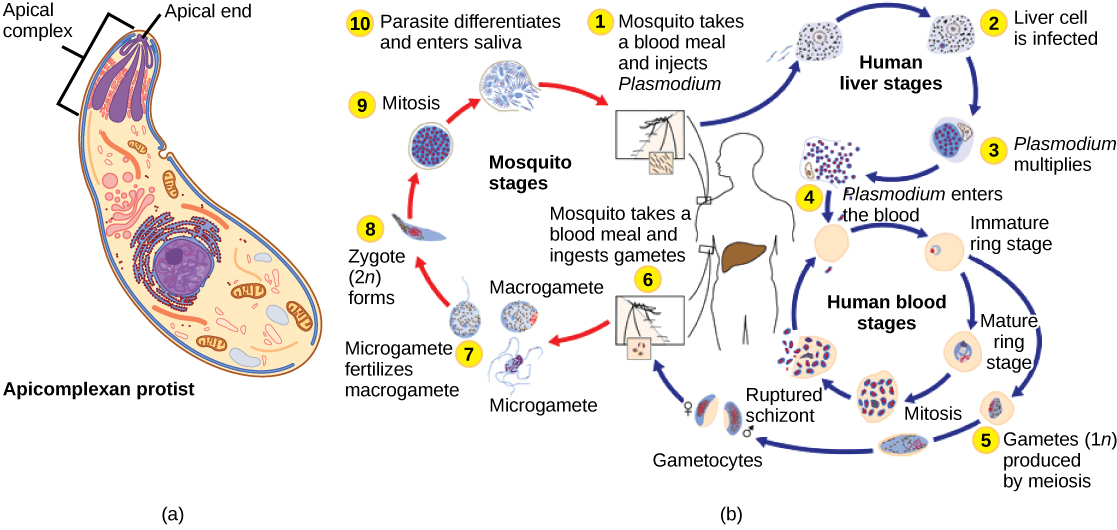
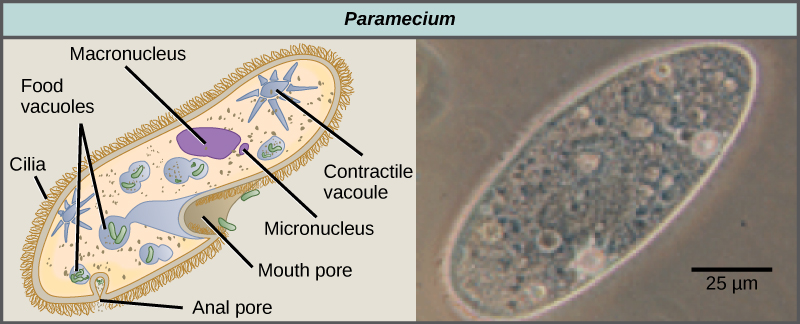
Los ciliados, que incluyen Paramecio y Tetrahymena, son un grupo de protistas de 10 a 3,000 micrómetros de longitud que están cubiertos de hileras, mechones o espirales de cilios diminutos. Al golpear sus cilios sincrónicamente o en ondas, los ciliados pueden coordinar movimientos dirigidos e ingerir partículas de alimentos. Ciertos ciliados tienen estructuras fundidas basadas en cilias que funcionan como paletas, embudos o aletas. Los ciliados también están rodeados por una película, proporcionando protección sin comprometer la agilidad. El género Paramecium incluye protistas que han organizado sus cilios en una boca primitiva en forma de placa, llamada surco oral, que se utiliza para capturar y digerir bacterias (Figura\(\PageIndex{7}\)). Los alimentos capturados en el surco oral entran en una vacuola alimentaria, donde se combina con enzimas digestivas. Las partículas residuales son expulsadas por una vesícula exocítica que se fusiona en una región específica de la membrana celular, llamada poro anal. Además de un sistema digestivo basado en vacuolas, Paramecium también utiliza vacuolas contráctiles, que son vesículas osmorreguladoras que se llenan de agua a medida que ingresa a la célula por ósmosis y luego se contraen para exprimir el agua de la célula.
Enlace al aprendizaje
Mira el video de la vacuola contráctil de Paramecio expulsando agua para mantener la célula osmóticamente equilibrada.
Paramecio tiene dos núcleos, un macronúcleo y un micronúcleo, en cada célula. El micronúcleo es esencial para la reproducción sexual, mientras que el macronúcleo dirige la fisión binaria asexual y todas las demás funciones biológicas. El proceso de reproducción sexual en Paramecium subraya la importancia del micronúcleo para estos protistas. Paramecio y la mayoría de los otros ciliados se reproducen sexualmente por conjugación. Este proceso comienza cuando dos tipos de apareamiento diferentes de Paramecio hacen contacto físico y se unen con un puente citoplásmico (Figura\(\PageIndex{8}\)). El micronúcleo diploide en cada célula se somete a meiosis para producir cuatro micronúcleos haploides. Tres de estos degeneran en cada célula, dejando un micronúcleo que luego sufre mitosis, generando dos micronúcleos haploides. Cada una de las células intercambia uno de estos núcleos haploides y se alejan unas de otras. Un proceso similar ocurre en bacterias que tienen plásmidos. La fusión de los micronúcleos haploides genera un pre-micronúcleo diploide completamente novedoso en cada célula conjugativa. Este pre-micronúcleo se somete a tres rondas de mitosis para producir ocho copias, y el macronúcleo original se desintegra. Cuatro de los ocho pre-micronúcleos se convierten en micronúcleos completos, mientras que los otros cuatro realizan múltiples rondas de replicación del ADN y se convierten en nuevos macronúcleos. Dos divisiones celulares producen cuatro nuevas Paramecia a partir de cada célula conjugativa original.
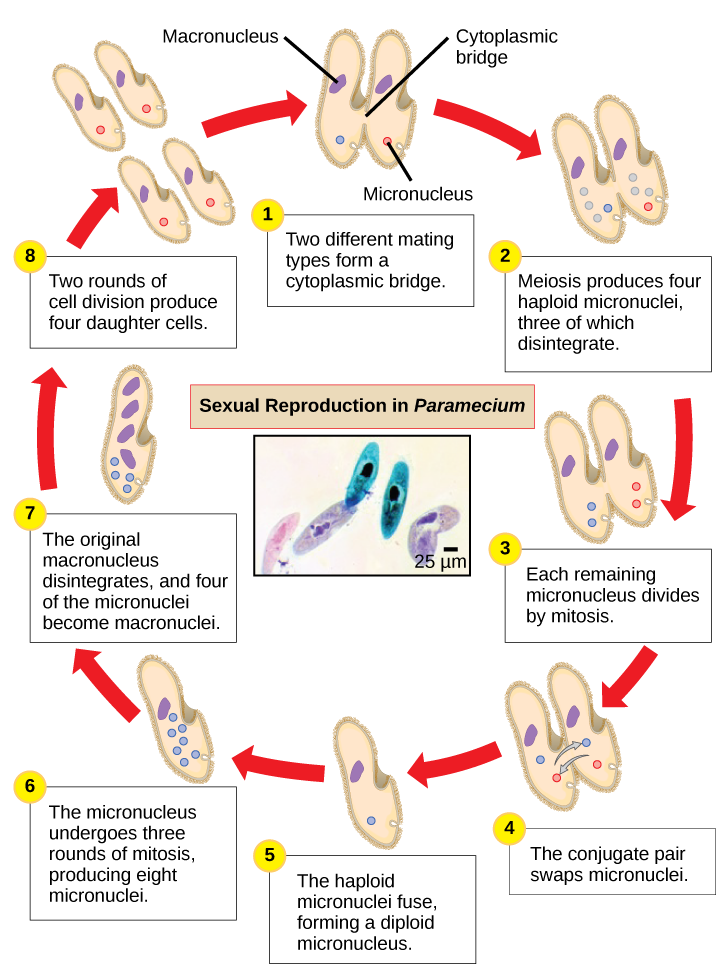
Ejercicio
¿Cuál de las siguientes afirmaciones sobre la reproducción sexual de Paramecium es falsa?
- Los macronúcleos se derivan de micronúcleos.
- Tanto la mitosis como la meiosis ocurren durante la reproducción sexual.
- El par conjugado intercambia macronúcleos.
- Cada padre produce cuatro células hijas.
Estramenopiles: Diatomeas, Algas Pardas, Algas Doradas y Oomicetos
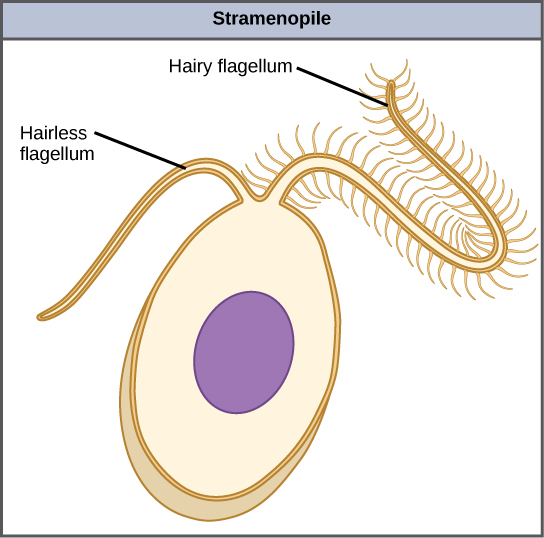
El otro subgrupo de cromalveolatos, los estramenopiles, incluye algas marinas fotosintéticas y protistas heterótrofos. La característica unificadora de este grupo es la presencia de un flagelo texturizado o “peludo”. Muchos estramenopiles también tienen un flagelo adicional que carece de proyecciones en forma de pelo (Figura\(\PageIndex{9}\)). Los miembros de este subgrupo varían en tamaño desde diatomeas unicelulares hasta las algas masivas y multicelulares.
Las diatomeas son protistas fotosintéticos unicelulares que se encierran en paredes celulares vítreas y intrincadamente estampadas compuestas por dióxido de silicio en una matriz de partículas orgánicas (Figura\(\PageIndex{10}\)). Estos protistas son un componente del plancton de agua dulce y marino. La mayoría de las especies de diatomeas se reproducen asexualmente, aunque también existen algunos casos de reproducción sexual y esporulación. Algunas diatomeas exhiben una hendidura en su cubierta de sílice, llamada rafe. Al expulsar una corriente de mucopolisacáridos del rafe, la diatomea puede unirse a las superficies o impulsarse en una dirección.
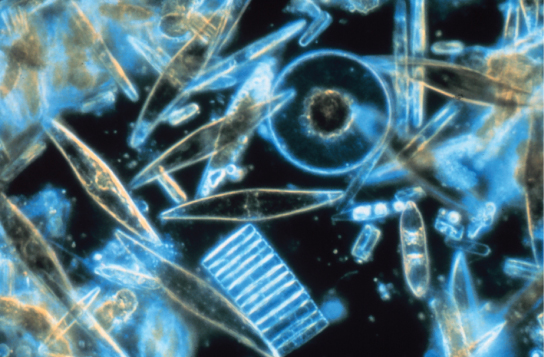
Durante los períodos de disponibilidad de nutrientes, las poblaciones de diatomeas florecen a números mayores que los que pueden consumir los organismos acuáticos. El exceso de diatomeas muere y se hunde hasta el fondo marino donde no son fácilmente alcanzadas por sasondas que se alimentan de organismos muertos. Como resultado, el dióxido de carbono que las diatomeas habían consumido e incorporado a sus células durante la fotosíntesis no se devuelve a la atmósfera. En general, este proceso por el cual el carbono es transportado profundamente al océano se describe como la bomba de carbono biológico, debido a que el carbono es “bombeado” a las profundidades oceánicas donde es inaccesible a la atmósfera como dióxido de carbono. La bomba de carbono biológico es un componente crucial del ciclo del carbono que mantiene niveles más bajos de dióxido de carbono atmosférico.
Al igual que las diatomeas, las algas doradas son en gran parte unicelulares, aunque algunas especies pueden formar colonias Su característico color dorado es el resultado de su uso extensivo de carotenoides, un grupo de pigmentos fotosintéticos que generalmente son de color amarillo o naranja. Las algas doradas se encuentran tanto en ambientes de agua dulce como marinos, donde forman una parte importante de la comunidad de plancton.
Las algas pardas son principalmente organismos marinos, multicelulares que se conocen coloquialmente como algas marinas. Las algas gigantes son un tipo de algas pardas. Algunas algas pardas han desarrollado tejidos especializados que se asemejan a las plantas terrestres, con retenes similares a las raíces, estipes similares a tallos y hojas en forma de hojas que son capaces de fotosíntesis. Los stipes de las algas gigantes son enormes, extendiéndose en algunos casos por 60 metros. Existe una variedad de ciclos de vida de algas, pero el más complejo es la alternancia de generaciones, en las que tanto la etapa haploide como la diploide implican multicelularidad. Compara este ciclo de vida con el de los humanos, por ejemplo. Los gametos haploides producidos por la meiosis (esperma y óvulo) se combinan en la fertilización para generar un cigoto diploide que sufre muchas rondas de mitosis para producir un embrión multicelular y luego un feto. Sin embargo, los espermatozoides y óvulos individuales nunca se convierten en seres multicelulares. Las plantas terrestres también han evolucionado alternancia de generaciones. En el género de algas pardas Laminaria, las esporas haploides se desarrollan en gametofitos multicelulares, los cuales producen gametos haploides que se combinan para producir organismos diploides que luego se convierten en organismos multicelulares con una estructura diferente a la forma haploide (Figura\(\PageIndex{11}\)). Algunos otros organismos realizan alternancia de generaciones en las que tanto la forma haploide como la diploide tienen el mismo aspecto.
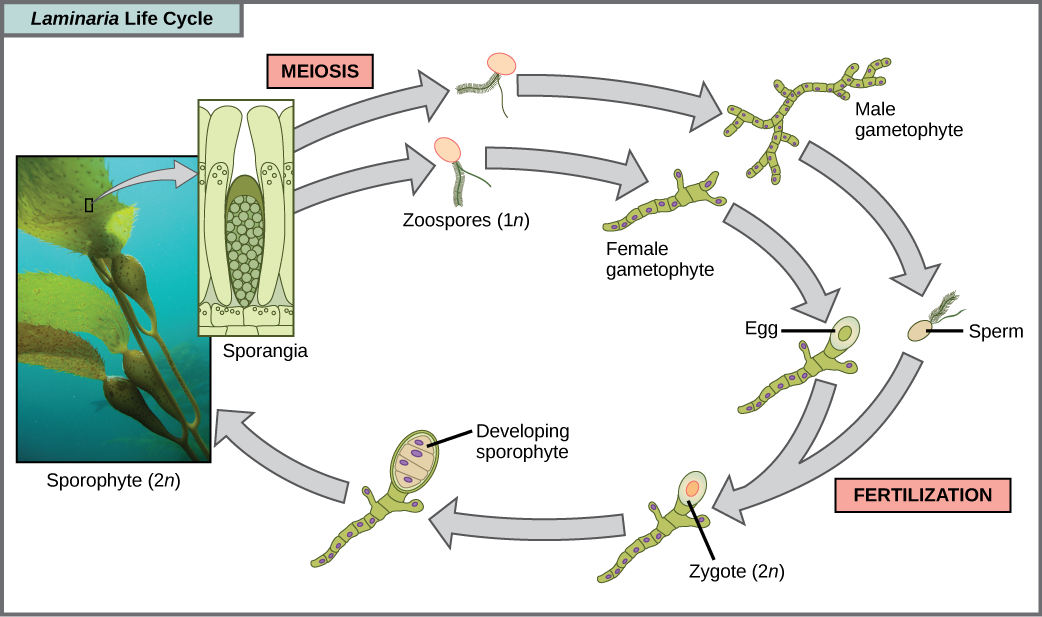
Ejercicio
¿Cuál de las siguientes afirmaciones sobre el ciclo de vida de Laminaria es falsa?
- Se forman 1 n zoosporas en los esporangios.
- El esporófito es la planta de 2 n.
- El gametofito es diploide.
- Tanto los estadios gametófitos como los esporófitos son multicelulares.
Los mohos de agua, oomicetos (“hongo del huevo”), fueron llamados por su morfología similar a hongos, pero los datos moleculares han demostrado que los mohos de agua no están estrechamente relacionados con los hongos. Los oomicetos se caracterizan por una pared celular a base de celulosa y una extensa red de filamentos que permiten la absorción de nutrientes. Como esporas diploides, muchos oomicetos tienen dos flagelos opuestos (uno peludo y otro liso) para la locomoción. Los oomicetos no son fotosintéticos e incluyen muchos sasondas y parásitos. Las sasondas aparecen como crecimientos esponjosos blancos en organismos muertos (Figura\(\PageIndex{12}\)). La mayoría de los oomicetos son acuáticos, pero algunos parasitan a las plantas terrestres. Un patógeno vegetal es Phytophthora infestans, el agente causante del tizón tardío de la papa, como ocurrió en la hambruna irlandesa de papa del siglo XIX.

Rizaria
El supergrupo Rhizaria incluye muchas de las amebas, la mayoría de las cuales tienen pseudopodios filiformes o agujereados (Figura\(\PageIndex{13}\)). Los pseudopodios funcionan para atrapar y engullir partículas de alimentos y dirigir el movimiento en protistas rizarios. Estos seudopodos se proyectan hacia afuera desde cualquier lugar de la superficie celular y pueden anclarse a un sustrato. El protista luego transporta su citoplasma al pseudopodo, moviendo así toda la célula. Este tipo de movimiento, llamado streaming citoplásmico, es utilizado por varios grupos diversos de protistas como medio de locomoción o como método para distribuir nutrientes y oxígeno.
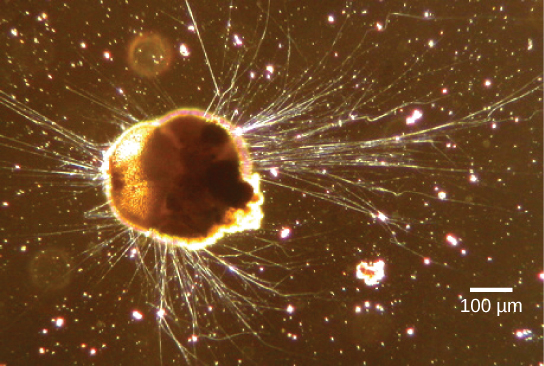
Enlace al aprendizaje
Echa un vistazo a este video para ver streaming citoplásmico en un alga verde.
Formas
Los foraminíferos, o forams, son protistas heterótrofos unicelulares, que van desde aproximadamente 20 micrómetros hasta varios centímetros de longitud, y ocasionalmente se asemejan a caracoles diminutos (Figura\(\PageIndex{14}\)). Como grupo, los forams exhiben conchas porosas, llamadas pruebas que se construyen a partir de diversos materiales orgánicos y típicamente endurecidos con carbonato de calcio. Las pruebas pueden albergar algas fotosintéticas, que los forams pueden cosechar para su nutrición. Los pseudopodios de foram se extienden a través de los poros y permiten que los forams se muevan, alimenten y reúnan materiales de construcción adicionales. Por lo general, las foreras están asociadas con arena u otras partículas en hábitats marinos o de agua dulce. Los foraminíferos también son útiles como indicadores de contaminación y cambios en los patrones climáticos globales.

Radiolarios
Un segundo subtipo de Rhizaria, los radiolarios, exhiben intrincados exteriores de sílice vítreo con simetría radial o bilateral (Figura\(\PageIndex{15}\)). Los pseudopodos en forma de aguja soportados por microtúbulos irradian hacia afuera desde los cuerpos celulares de estos protistas y funcionan para atrapar partículas de alimentos. Las conchas de radiolarios muertos se hunden hasta el fondo del océano, donde pueden acumularse en profundidades de 100 metros de espesor. Los radiolarios conservados y sedimentados son muy comunes en el registro fósil.
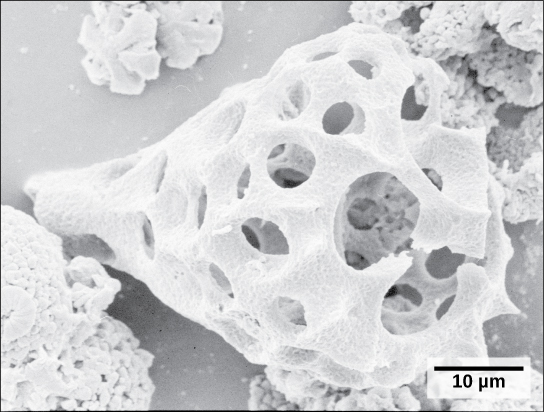
Archaeplastida
Las algas rojas y verdes están incluidas en el supergrupo Archaeplastida. Fue a partir de un ancestro común de estos protistas que las plantas terrestres evolucionaron, ya que sus parientes más cercanos se encuentran en este grupo. La evidencia molecular sostiene que todos los Archaeplastida son descendientes de una relación endosimbiótica entre un protista heterotrófico y una cianobacteria. Las algas rojas y verdes incluyen formas unicelulares, multicelulares y coloniales.
Algas Rojas
Las algas rojas, o rodófitas, son principalmente multicelulares, carecen de flagelos y varían en tamaño desde protistas microscópicos unicelulares hasta formas multicelulares grandes agrupadas en la categoría de algas informales. El ciclo de vida de las algas rojas es una alternancia de generaciones. Algunas especies de algas rojas contienen ficoeritrinas, pigmentos accesorios fotosintéticos que son de color rojo y superan al tinte verde de la clorofila, haciendo que estas especies aparezcan como diferentes tonos de rojo. Otros protistas clasificados como algas rojas carecen de ficoeritrinas y son parásitos. Las algas rojas son comunes en aguas tropicales donde se han detectado a profundidades de 260 metros. Otras algas rojas existen en ambientes terrestres o de agua dulce.
Algas Verdes: Clorofitos y Carófitos
El grupo de algas más abundante son las algas verdes. Las algas verdes presentan características similares a las plantas terrestres, particularmente en términos de estructura de cloroplastos. El hecho de que este grupo de protistas compartiera un ancestro común relativamente reciente con las plantas terrestres está bien sustentado. Las algas verdes se subdividen en las clorofitas y las carófitas. Los carófitos son los parientes vivos más cercanos a las plantas terrestres y se asemejan a ellos en morfología y estrategias reproductivas. Los charófitos son comunes en hábitats húmedos, y su presencia a menudo indica un ecosistema saludable.
Los clorofitos presentan gran diversidad de forma y función. Los clorofitos habitan principalmente el suelo de agua dulce y húmedo, y son un componente común del plancton. La clamidomonas es una clorofita simple y unicelular con una morfología en forma de pera y dos flagelos anteriores opuestos que guían a este protista hacia la luz percibida por su mancha ocular. Las especies de clorofito más complejas exhiben gametos haploides y esporas que se asemejan a Chlamydomonas.
El clorofito Volvox es uno de los pocos ejemplos de un organismo colonial, que se comporta de alguna manera como una colección de células individuales, pero de otras formas como las células especializadas de un organismo multicelular (Figura\(\PageIndex{16}\)). Las colonias Volvox contienen de 500 a 60,000 células, cada una con dos flagelos, contenidas dentro de una matriz hueca y esférica compuesta por una secreción de glicoproteína gelatinosa. Las células Volvox individuales se mueven de manera coordinada y están interconectadas por puentes citoplásmicos. Sólo algunas de las células se reproducen para crear colonias hijas, ejemplo de especialización celular básica en este organismo.
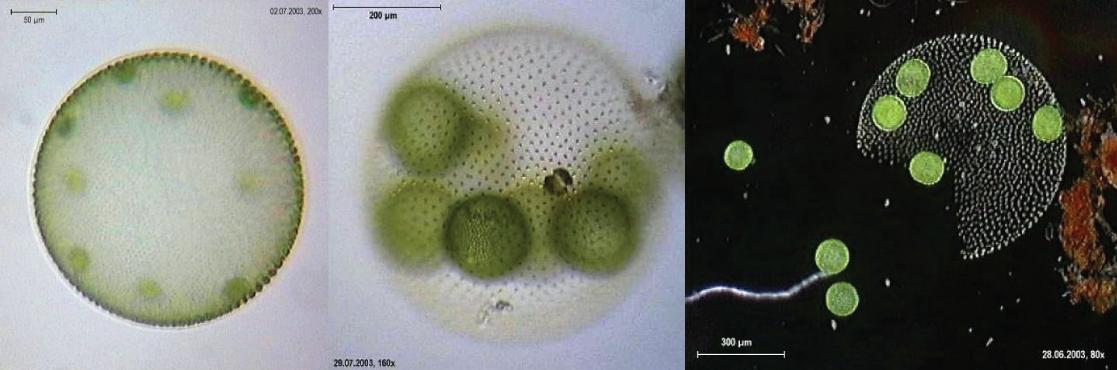
Verdaderos organismos multicelulares, como la lechuga de mar, Ulva, están representados entre las clorofitas. Además, algunas clorofitas existen como células grandes, multinucleadas, individuales. Las especies del género Caulerpa exhiben follaje aplanado similar a helecho y pueden alcanzar longitudes de 3 metros (Figura\(\PageIndex{17}\)). Las especies de Caulerpa experimentan división nuclear, pero sus células no completan la citocinesis, permaneciendo en su lugar como células individuales masivas y elaboradas.

Amoebozoa
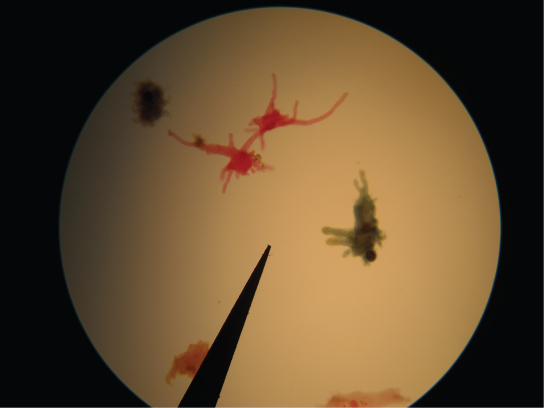
Los amebozoos presentan característicamente pseudopodios que se extienden como tubos o lóbulos planos, en lugar de los pseudopodios similares a pelo de la ameba rizaria (Figura\(\PageIndex{18}\)). Los amebozoos incluyen varios grupos de organismos unicelulares similares a las amebas que son de vida libre o parásitos.
Moldes de limo
Un subconjunto de los amebozoos, los mohos de limo, tiene varias similitudes morfológicas con los hongos que se cree que son el resultado de la evolución convergente. Por ejemplo, en momentos de estrés, algunos mohos de limo se convierten en cuerpos fructíferos generadores de esporas, al igual que los hongos.
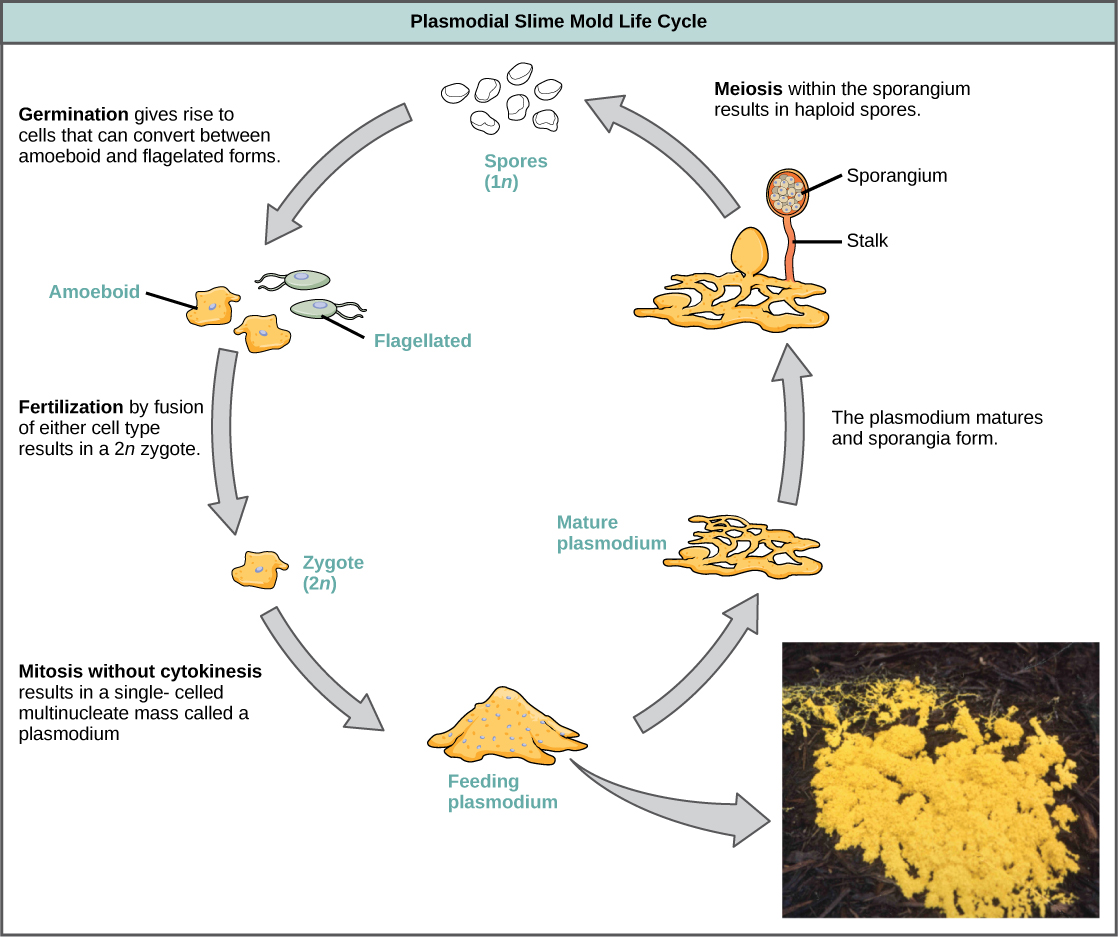
Los mohos de limo se categorizan en función de sus ciclos de vida en tipos plasmodiales o celulares. Los mohos plasmodiales están compuestos por células grandes multinucleadas y se mueven a lo largo de las superficies como una gota amorfa de limo durante su etapa de alimentación (Figura\(\PageIndex{19}\)). Las partículas de alimentos se levantan y se envuelven en el molde de limo a medida que se desliza. Al madurar, el plasmodio adquiere una apariencia de red con la capacidad de formar cuerpos fructíferos, o esporangios, en momentos de estrés. Las esporas haploides son producidas por la meiosis dentro de los esporangios, y las esporas pueden diseminarse a través del aire o el agua para aterrizar potencialmente en ambientes más favorables. Si esto ocurre, las esporas germinan para formar células haploides ameboides o flageladas que pueden combinarse entre sí y producir un moho cigótico diploide de limo para completar el ciclo de vida.
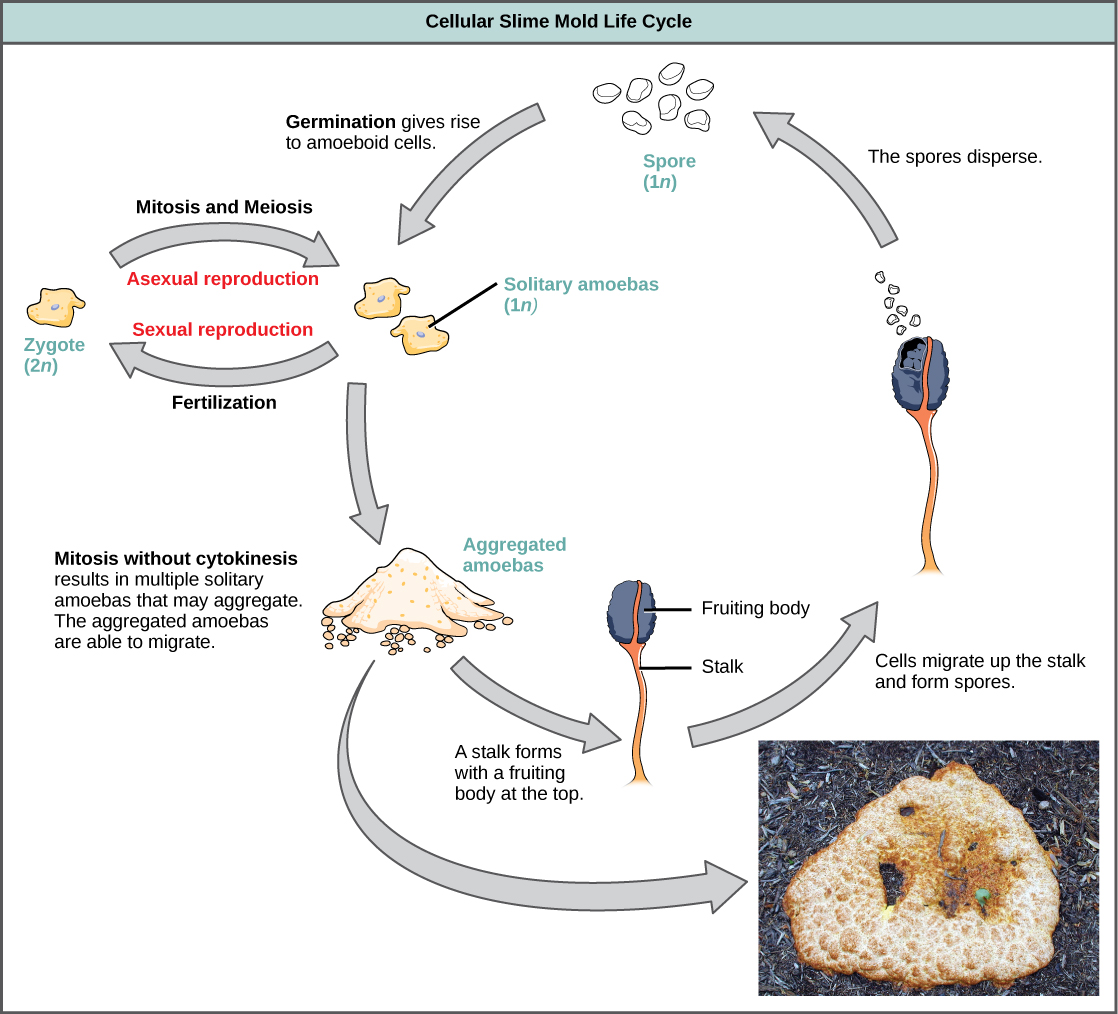
Los mohos celulares funcionan como células ameboides independientes cuando abundan los nutrientes (Figura\(\PageIndex{20}\)). Cuando los alimentos se agotan, los mohos celulares de limo se apilan entre sí en una masa de celdas que se comporta como una sola unidad, llamada babosa. Algunas células en la babosa contribuyen a un tallo de 2-3 milímetros, secándose y muriendo en el proceso. Las células en la parte superior del tallo forman un cuerpo fructífero asexual que contiene esporas haploides. Al igual que con los mohos plasmodiales, las esporas se diseminan y pueden germinar si aterrizan en un ambiente húmedo. Un género representativo de los mohos celulares de limo es Dictyostelium, que comúnmente existe en el suelo húmedo de los bosques.
Enlace al aprendizaje
Visualiza este sitio para ver la formación de un cuerpo fructífero por un moho celular de limo.
Opisthokonta
Los opisthokonts incluyen los choanoflagelados similares a animales, que se cree que se asemejan al ancestro común de las esponjas y, de hecho, a todos los animales. Los choanoflagelados incluyen formas unicelulares y coloniales, y suman alrededor de 244 especies descritas. Estos organismos exhiben un solo flagelo apical que está rodeado por un collar contráctil compuesto por microvellosidades. El collar utiliza un mecanismo similar a las esponjas para filtrar las bacterias para su ingestión por el protista. La morfología de los coanoflagelados se reconoció desde el principio como parecida a las células del collar de las esponjas, y sugiriendo una posible relación con los animales.
Los Mesomicetozoos forman un pequeño grupo de parásitos, principalmente de peces, y al menos una forma que puede parasitar a los humanos. Sus ciclos de vida son poco comprendidos. Estos organismos son de especial interés, porque parecen estar tan estrechamente relacionados con los animales. En el pasado, se agruparon con hongos y otros protistas en función de su morfología.
Resumen
El proceso de clasificar a los protistas en grupos significativos está en curso, pero los datos genéticos en los últimos 20 años han aclarado muchas relaciones que antes no estaban claras o equivocadas. La opinión mayoritaria en la actualidad es ordenar a todos los eucariotas en seis supergrupos: Excavata, Chromalveolata, Rhizaria, Archaeplastida, Amoebozoa y Opisthokonta. El objetivo de este esquema de clasificación es crear cúmulos de especies que todas deriven de un ancestro común. En la actualidad, la monofilia de algunos de los supergrupos está mejor sustentada por datos genéticos que otros. Aunque existe una tremenda variación dentro de los supergrupos, se pueden identificar puntos en común a nivel morfológico, fisiológico y ecológico.
Conexiones de arte
Figura\(\PageIndex{8}\): ¿Cuál de las siguientes afirmaciones sobre la reproducción sexual de Paramecium es falsa?
- Los macronúcleos se derivan de micronúcleos.
- Tanto la mitosis como la meiosis ocurren durante la reproducción sexual.
- El par conjugado intercambia macronúcleos.
- Cada padre produce cuatro células hijas.
- Contestar
-
C
Figura\(\PageIndex{11}\): ¿Cuál de las siguientes afirmaciones sobre el ciclo de vida de Laminaria es falsa?
- Se forman 1 n zoosporas en los esporangios.
- El esporófito es la planta de 2 n.
- El gametofito es diploide.
- Tanto los estadios gametófitos como los esporófitos son multicelulares.
- Contestar
-
C
Glosario
- bomba de carbono biológico
- proceso por el cual el carbono inorgánico es fijado por especies fotosintéticas que luego mueren y caen al fondo marino donde no pueden ser alcanzados por saprobes y su consumo de dióxido de carbono no puede ser devuelto a la atmósfera
- bioluminiscencia
- generación y emisión de luz por un organismo, como en dinoflagelados
- vacuola contráctil
- vesícula que se llena de agua (cuando entra en la célula por ósmosis) y luego se contrae para exprimir el agua de la célula; una vesícula osmorreguladora
- streaming citoplásmico
- movimiento del citoplasma en un pseudopodo extendido de tal manera que toda la célula es transportada al sitio del pseudopodo
- hidrogenosoma
- organelo portado por parabasálidos (Excavata) que funciona anaeróbicamente y produce gas hidrógeno como subproducto; probablemente evolucionado a partir de mitocondrias
- kinetoplasto
- masa de ADN transportada dentro de la mitocondria única sobredimensionada, característica de cinetoplastos (filo: Euglenozoa)
- mitosoma
- orgánulos no funcionales transportados en las células de diplomonadas (Excavata) que probablemente evolucionaron a partir de una mitocondria
- plancton
- grupo diverso de organismos en su mayoría microscópicos que se desplazan en sistemas marinos y de agua dulce y sirven como fuente de alimento para organismos acuáticos más grandes
- raphe
- hendidura en la cubierta de sílice de diatomeas a través de la cual el protista secreta una corriente de mucopolisacáridos para locomoción y unión a sustratos
- prueba
- cáscara porosa de una forma que está construida a partir de diversos materiales orgánicos y típicamente endurecida con carbonato de calcio