
4.3: Ciclo del ácido cítrico y fosforilación oxidativa
- Page ID
- 54364

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)El ciclo del ácido cítrico
En las células eucariotas, las moléculas de piruvato producidas al final de la glucólisis son transportadas a las mitocondrias, que son sitios de respiración celular. Si hay oxígeno disponible, la respiración aeróbica seguirá adelante. En las mitocondrias, el piruvato se transformará en un grupo acetilo de dos carbonos (mediante la eliminación de una molécula de dióxido de carbono) que será captado por un compuesto portador llamado coenzima A (CoA), que se elabora a partir de la vitamina B 5. El compuesto resultante se llama acetil CoA. (Figura\(\PageIndex{1}\)). El acetil CoA puede ser utilizado de diversas maneras por la célula, pero su función principal es entregar el grupo acetilo derivado del piruvato a la siguiente vía en el catabolismo de la glucosa.
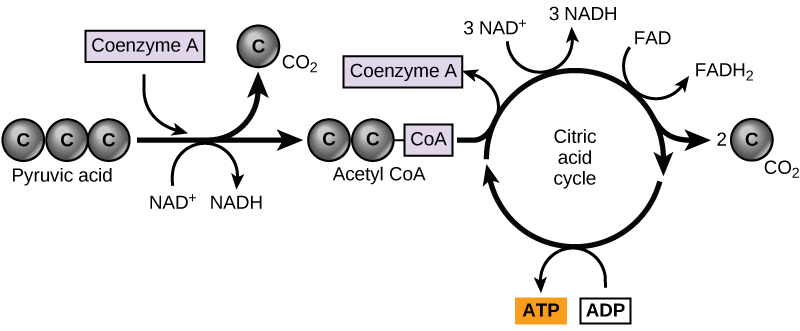
Al igual que la conversión del piruvato en acetil CoA, el ciclo del ácido cítrico en las células eucariotas tiene lugar en la matriz de las mitocondrias. A diferencia de la glucólisis, el ciclo del ácido cítrico es un ciclo cerrado: La última parte de la vía regenera el compuesto utilizado en el primer paso. Los ocho pasos del ciclo son una serie de reacciones químicas que producen dos moléculas de dióxido de carbono, una molécula de ATP (o una equivalente), y formas reducidas (NADH y FADH 2) de NAD + y FAD +, coenzimas importantes en la célula. Parte de esto se considera una vía aeróbica (que requiere oxígeno) porque el NADH y el FADH 2 producidos deben transferir sus electrones a la siguiente vía en el sistema, que utilizará oxígeno. Si el oxígeno no está presente, esta transferencia no ocurre.
Dos átomos de carbono entran en el ciclo del ácido cítrico de cada grupo acetilo. Dos moléculas de dióxido de carbono se liberan en cada giro del ciclo; sin embargo, estas no contienen los mismos átomos de carbono aportados por el grupo acetilo en ese giro de la ruta. Los dos átomos de acetil-carbono eventualmente se liberarán en giros posteriores del ciclo; de esta manera, los seis átomos de carbono de la molécula de glucosa original se liberarán eventualmente como dióxido de carbono. Se necesitan dos vueltas del ciclo para procesar el equivalente de una molécula de glucosa. Cada giro del ciclo forma tres moléculas de NADH de alta energía y una molécula FADH 2 de alta energía. Estos portadores de alta energía se conectarán con la última porción de la respiración aeróbica para producir moléculas de ATP. También se hace un ATP (o un equivalente) en cada ciclo. Varios de los compuestos intermedios en el ciclo del ácido cítrico pueden ser utilizados en la síntesis de aminoácidos no esenciales; por lo tanto, el ciclo es tanto anabólico como catabólico.
Fosforilación oxidativa
Acabas de leer sobre dos vías en el catabolismo de la glucosa, la glucólisis y el ciclo del ácido cítrico, que generan ATP. La mayor parte del ATP generado durante el catabolismo aeróbico de la glucosa, sin embargo, no se genera directamente a partir de estas vías. Más bien, deriva de un proceso que comienza con el paso de electrones a través de una serie de reacciones químicas a un aceptor de electrones final, el oxígeno. Estas reacciones tienen lugar en complejos proteicos especializados ubicados en la membrana interna de las mitocondrias de organismos eucariotas y en la parte interna de la membrana celular de organismos procariotas. La energía de los electrones es cosechada y utilizada para generar un gradiente electroquímico a través de la membrana mitocondrial interna. La energía potencial de este gradiente se utiliza para generar ATP. La totalidad de este proceso se denomina fosforilación oxidativa.
La cadena de transporte de electrones (Figura\(\PageIndex{2}\) a) es el último componente de la respiración aeróbica y es la única parte del metabolismo que utiliza oxígeno atmosférico. El oxígeno se difunde continuamente en las plantas para este propósito. En los animales, el oxígeno ingresa al cuerpo a través del sistema respiratorio. El transporte de electrones es una serie de reacciones químicas que se asemejan a una brigada de baldes en que los electrones pasan rápidamente de un componente al siguiente, al punto final de la cadena donde el oxígeno es el aceptor de electrones final y se produce agua. Hay cuatro complejos compuestos por proteínas, etiquetados de I a IV en la Figura\(\PageIndex{2}\) c, y la agregación de estos cuatro complejos, junto con los portadores de electrones móviles asociados, se llama la cadena de transporte de electrones. La cadena de transporte de electrones está presente en múltiples copias en la membrana mitocondrial interna de eucariotas y en la membrana plasmática de procariotas. En cada transferencia de un electrón a través de la cadena de transporte de electrones, el electrón pierde energía, pero con algunas transferencias, la energía se almacena como energía potencial utilizándola para bombear iones de hidrógeno a través de la membrana mitocondrial interna hacia el espacio intermembrana, creando un gradiente electroquímico.
CONEXIÓN ARTE
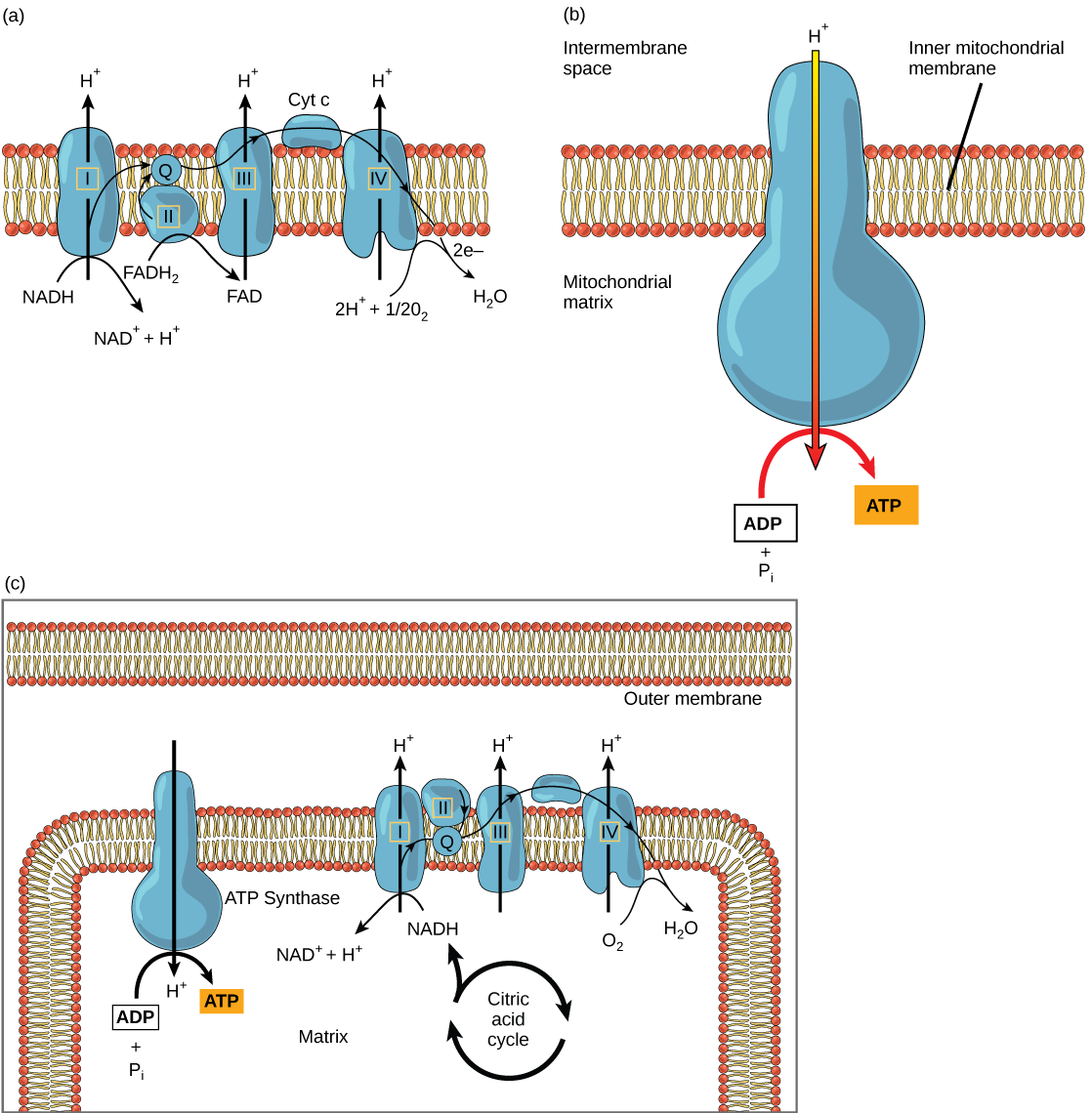
El cianuro inhibe la citocromo c oxidasa, un componente de la cadena de transporte de electrones. Si se produce envenenamiento por cianuro, ¿esperaría que el pH del espacio intermembrana aumente o disminuya? ¿Qué efecto tendría el cianuro en la síntesis de ATP?
Los electrones de NADH y FADH 2 se pasan a complejos proteicos en la cadena de transporte de electrones. A medida que pasan de un complejo a otro (hay un total de cuatro), los electrones pierden energía, y parte de esa energía se utiliza para bombear iones de hidrógeno desde la matriz mitocondrial al espacio intermembrana. En el cuarto complejo proteico, los electrones son aceptados por el oxígeno, el aceptor terminal. El oxígeno con sus electrones adicionales se combina luego con dos iones de hidrógeno, mejorando aún más el gradiente electroquímico, para formar agua. Si no hubiera oxígeno presente en la mitocondria, los electrones no podrían eliminarse del sistema, y toda la cadena de transporte de electrones retrocedería y se detendría. Las mitocondrias serían incapaces de generar nuevos ATP de esta manera, y la célula moriría en última instancia por falta de energía. Esta es la razón por la que debemos respirar para extraer nuevo oxígeno.
En la cadena de transporte de electrones, la energía libre de la serie de reacciones que se acaban de describir se utiliza para bombear iones de hidrógeno a través de la membrana. La distribución desigual de iones H + a través de la membrana establece un gradiente electroquímico, debido a la carga positiva de los iones H + y su mayor concentración en un lado de la membrana.
Los iones de hidrógeno se difunden a través de la membrana interna a través de una proteína integral de membrana llamada ATP sintasa (Figura\(\PageIndex{2}\) b). Esta compleja proteína actúa como un pequeño generador, girado por la fuerza de los iones de hidrógeno que se difunden a través de ella, bajando por su gradiente electroquímico desde el espacio intermembrana, donde hay muchos iones de hidrógeno que se repelen mutuamente a la matriz, donde hay pocos. El giro de las partes de esta máquina molecular regenera ATP a partir de ADP. Este flujo de iones de hidrógeno a través de la membrana a través de la ATP sintasa se llama quimiósmosis.
La quimiósmosis (Figura\(\PageIndex{2}\) c) se utiliza para generar 90 por ciento del ATP producido durante el catabolismo de glucosa aeróbica. El resultado de las reacciones es la producción de ATP a partir de la energía de los electrones retirados de los átomos de hidrógeno. Estos átomos eran originalmente parte de una molécula de glucosa. Al final del sistema de transporte de electrones, los electrones se utilizan para reducir una molécula de oxígeno a iones de oxígeno. Los electrones adicionales en los iones de oxígeno atraen iones de hidrógeno (protones) del medio circundante, y se forma agua. La cadena de transporte de electrones y la producción de ATP a través de la quimiósmosis se denominan colectivamente fosforilación oxidativa.
Rendimiento ATP
El número de moléculas de ATP generadas a partir del catabolismo de la glucosa varía. Por ejemplo, el número de iones hidrógeno que los complejos de la cadena de transporte de electrones pueden bombear a través de la membrana varía entre especies. Otra fuente de varianza proviene de la lanzadera de electrones a través de la membrana mitocondrial. El NADH generado a partir de la glucólisis no puede ingresar fácilmente a las mitocondrias. Así, los electrones son captados en el interior de las mitocondrias por NAD + o FAD +. Se generan menos moléculas de ATP cuando FAD+ actúa como portador. El NAD + se utiliza como transportador de electrones en el hígado y FAD+ en el cerebro, por lo que el rendimiento de ATP depende del tejido que se esté considerando.
Otro factor que afecta el rendimiento de las moléculas de ATP generadas a partir de la glucosa es que los compuestos intermedios en estas vías se utilizan para otros fines. El catabolismo de la glucosa se conecta con las vías que construyen o descomponen todos los demás compuestos bioquímicos en las células, y el resultado es algo más desmesurado que las situaciones ideales descritas hasta ahora. Por ejemplo, los azúcares distintos de la glucosa se alimentan en la vía glucolítica para la extracción de energía. Otras moléculas que de otro modo se usarían para cosechar energía en la glucólisis o el ciclo del ácido cítrico pueden eliminarse para formar ácidos nucleicos, aminoácidos, lípidos u otros compuestos. En general, en los sistemas vivos, estas vías del catabolismo de la glucosa extraen alrededor del 34 por ciento de la energía contenida en la glucosa.
CARRERAS EN ACCIÓN: Médico de Enfermedades Mitocondriales
¿Qué sucede cuando las reacciones críticas de la respiración celular no proceden correctamente? Las enfermedades mitocondriales son trastornos genéticos del metabolismo. Los trastornos mitocondriales pueden surgir de mutaciones en el ADN nuclear o mitocondrial, y resultan en la producción de menos energía de lo normal en las células del cuerpo. Los síntomas de las enfermedades mitocondriales pueden incluir debilidad muscular, falta de coordinación, episodios similares a accidentes cerebrovasculares y pérdida de visión y audición. La mayoría de las personas afectadas son diagnosticadas en la infancia, aunque existen algunas enfermedades de inicio en la edad adulta. Identificar y tratar los trastornos mitocondriales es un campo médico especializado. La preparación educativa para esta profesión requiere de una educación universitaria, seguida de la escuela de medicina con especialización en genética médica. Los genetistas médicos pueden ser certificados por la Junta Americana de Genética Médica y llegar a asociarse con organizaciones profesionales dedicadas al estudio de la enfermedad mitocondrial, como la Sociedad de Medicina Mitocondrial y la Sociedad para la Enfermedad Metabólica Heredada.
Resumen
El ciclo del ácido cítrico es una serie de reacciones químicas que elimina electrones de alta energía y los utiliza en la cadena de transporte de electrones para generar ATP. Se produce una molécula de ATP (o un equivalente) por cada giro del ciclo.
La cadena de transporte de electrones es la porción de respiración aeróbica que utiliza oxígeno libre como aceptor final de electrones para los electrones eliminados de los compuestos intermedios en el catabolismo de glucosa. Los electrones pasan a través de una serie de reacciones químicas, con una pequeña cantidad de energía libre utilizada en tres puntos para transportar iones de hidrógeno a través de la membrana. Esto contribuye al gradiente utilizado en la quimiósmosis. A medida que los electrones pasan del NADH o FADH 2 por la cadena de transporte de electrones, pierden energía. Los productos de la cadena de transporte de electrones son el agua y el ATP. Varios compuestos intermedios pueden desviarse hacia el anabolismo de otras moléculas bioquímicas, tales como ácidos nucleicos, aminoácidos no esenciales, azúcares y lípidos. Estas mismas moléculas, excepto los ácidos nucleicos, pueden servir como fuentes de energía para la vía de la glucosa.
Conexiones de arte
Figura\(\PageIndex{2}\): El cianuro inhibe la citocromo c oxidasa, un componente de la cadena de transporte de electrones. Si se produce envenenamiento por cianuro, ¿esperaría que el pH del espacio intermembrana aumente o disminuya? ¿Qué efecto tendría el cianuro en la síntesis de ATP?
- Contestar
-
Después del envenenamiento por cianuro, la cadena de transporte de electrones ya no puede bombear electrones al espacio intermembrana. El pH del espacio intermembrana aumentaría y la síntesis de ATP se detendría.
Glosario
- acetil CoA
- la combinación de un grupo acetilo derivado del ácido pirúvico y la coenzima A que está hecha de ácido pantoténico (una vitamina del grupo B)
- ATP sintasa
- un complejo proteico embebido en la membrana que regenera ATP a partir de ADP con energía de protones que se difunden a través de él
- quimiósmosis
- el movimiento de iones de hidrógeno por su gradiente electroquímico a través de una membrana a través de ATP sintasa para generar ATP
- ciclo del ácido cítrico
- una serie de reacciones químicas catalizadas por enzimas de importancia central en todas las células vivas que cosechan la energía en enlaces carbono-carbono de las moléculas de azúcar para generar ATP; el ciclo del ácido cítrico es una vía metabólica aeróbica porque requiere oxígeno en reacciones posteriores para proceder
- cadena de transporte de electrones
- una serie de cuatro grandes complejos multiproteicos incrustados en la membrana mitocondrial interna que acepta electrones de compuestos donantes y recoge energía de una serie de reacciones químicas para generar un gradiente de iones de hidrógeno a través de la membrana
- fosforilación oxidativa
- la producción de ATP mediante la transferencia de electrones por la cadena de transporte de electrones para crear un gradiente de protones que es utilizado por la ATP sintasa para agregar grupos fosfato a las moléculas de ADP