
11.1: Descubriendo cómo cambian las poblaciones
- Page ID
- 53742

La teoría de la evolución por selección natural describe un mecanismo para el cambio de especies a lo largo del tiempo. Ese cambio de especie había sido sugerido y debatido mucho antes de Darwin. La visión de que las especies eran estáticas e inmutables se basaba en los escritos de Platón, sin embargo, también había antiguos griegos que expresaban ideas evolutivas.
En el siglo XVIII, las ideas sobre la evolución de los animales fueron reintroducidas por el naturalista Georges-Louis Leclerc, Comte de Buffon e incluso por el abuelo de Charles Darwin, Erasmus Darwin. Durante este tiempo, también se aceptó que había especies extintas. Al mismo tiempo, James Hutton, el naturalista escocés, propuso que el cambio geológico se producía gradualmente por la acumulación de pequeños cambios de procesos (a lo largo de largos periodos de tiempo) al igual que los que ocurren hoy en día. Esto contrastaba con la visión predominante de que la geología del planeta era consecuencia de eventos catastróficos ocurridos durante un pasado relativamente breve. La visión de Hutton fue posteriormente popularizada por el geólogo Charles Lyell en el siglo XIX. Lyell se hizo amigo de Darwin y sus ideas fueron muy influyentes en el pensamiento de Darwin. Lyell argumentó que la mayor edad de la Tierra dio más tiempo para el cambio gradual en las especies, y el proceso proporcionó una analogía para el cambio gradual en las especies.
A principios del siglo XIX, Jean-Baptiste Lamarck publicó un libro que detallaba un mecanismo para el cambio evolutivo que ahora se conoce como herencia de características adquiridas. En la teoría de Lamarck, las modificaciones en un individuo causadas por su entorno, o el uso o desuso de una estructura durante su vida, podrían ser heredadas por su descendencia y, así, provocar cambios en una especie. Si bien este mecanismo de cambio evolutivo descrito por Lamarck fue desacreditado, las ideas de Lamarck fueron una influencia importante en el pensamiento evolutivo. La inscripción en la estatua de Lamarck que se encuentra a las puertas del Jardin des Plantes en París lo describe como el “fundador de la doctrina de la evolución”.
Charles Darwin y la selección natural
El mecanismo real de evolución fue concebido y descrito independientemente por dos naturalistas, Charles Darwin y Alfred Russell Wallace, a mediados del siglo XIX. Es importante destacar que cada uno dedicó tiempo a explorar el mundo natural en expediciones a los trópicos. De 1831 a 1836, Darwin viajó por todo el mundo en H.M.S. Beagle, visitando América del Sur, Australia y el extremo sur de África. Wallace viajó a Brasil para recolectar insectos en la selva amazónica de 1848 a 1852 y al archipiélago malayo de 1854 a 1862. El viaje de Darwin, como los viajes posteriores de Wallace en el archipiélago malayo, incluyó paradas en varias cadenas de islas, siendo la última las Islas Galápagos (al oeste de Ecuador). En estas islas, Darwin observó especies de organismos en diferentes islas que eran claramente similares, pero que tenían diferencias distintas. Por ejemplo, los pinzones terrestres que habitaban las Islas Galápagos comprendían varias especies que cada una tenía una forma de pico única (Figura\(\PageIndex{1}\)). Observó tanto que estos pinzones se asemejaban mucho a otra especie de pinzón en el continente de Sudamérica como que el grupo de especies en las Galápagos formaba una serie graduada de tamaños y formas de pico, con diferencias muy pequeñas entre las más similares. Darwin imaginó que las especies de la isla podrían ser todas especies modificadas de una especie original del continente. En 1860, escribió: “Al ver esta gradación y diversidad de estructura en un pequeño grupo de aves íntimamente relacionado, uno realmente podría imaginarse que de una escasez original de aves en este archipiélago, una especie había sido tomada y modificada para diferentes fines”. 1
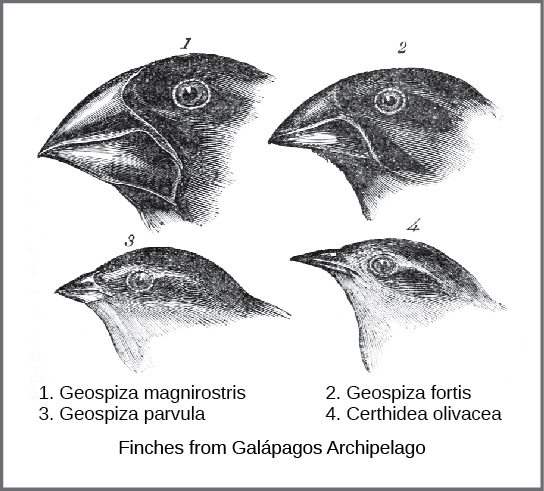
Wallace y Darwin observaron patrones similares en otros organismos e independientemente concibieron un mecanismo para explicar cómo y por qué podrían ocurrir tales cambios. Darwin llamó a este mecanismo de selección natural. La selección natural, argumentó Darwin, fue un resultado inevitable de tres principios que operaban en la naturaleza. En primer lugar, las características de los organismos se heredan, o se transmiten de padres a hijos. Segundo, se producen más crías de las que son capaces de sobrevivir; es decir, los recursos para la supervivencia y reproducción son limitados. La capacidad de reproducción en todos los organismos supera la disponibilidad de recursos para apoyar su número. Así, hay una competencia por esos recursos en cada generación. Tanto la comprensión de Darwin como Wallace de este principio provino de la lectura de un ensayo del economista Thomas Malthus, quien discutió este principio en relación con las poblaciones humanas. Tercero, las crías varían entre sí en cuanto a sus características y esas variaciones se heredan. De estos tres principios, Darwin y Wallace razonaron que las crías con características heredadas que les permitan competir mejor por recursos limitados sobrevivirán y tendrán más descendencia que aquellos individuos con variaciones que son menos capaces de competir. Debido a que las características son heredadas, estos rasgos estarán mejor representados en la próxima generación. Esto conducirá a un cambio en las poblaciones a lo largo de generaciones en un proceso que Darwin llamó “descenso con modificación”.
Los trabajos de Darwin y Wallace (Figura\(\PageIndex{2}\)) que presentan la idea de selección natural se leyeron juntos en 1858 antes de la Linnaean Society de Londres. Al año siguiente se publicó el libro de Darwin, Sobre el origen de las especies, que esbozó con considerable detalle sus argumentos a favor de la evolución por selección natural.
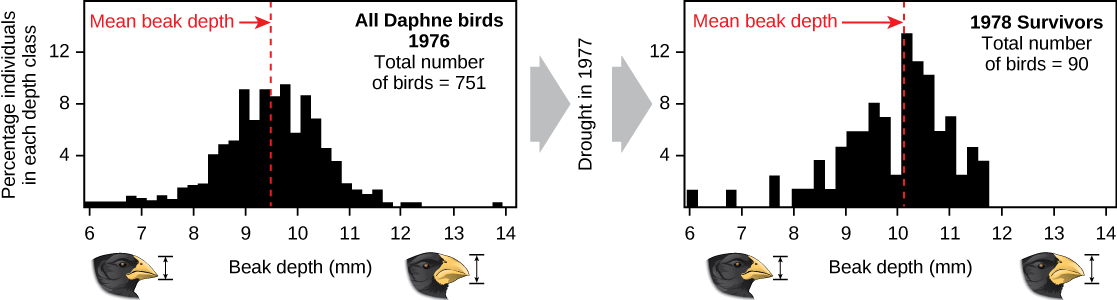
Las demostraciones de evolución por selección natural pueden llevar mucho tiempo. Una de las mejores demostraciones ha sido en las mismas aves que ayudaron a inspirar la teoría, los pinzones de Galápagos. Peter y Rosemary Grant y sus colegas han estudiado poblaciones de pinzones de Galápagos todos los años desde 1976 y han brindado importantes demostraciones del funcionamiento de la selección natural. Los Grants encontraron cambios de una generación a otra en las formas de pico de los pinzones de tierra mediana en la isla Galápagos de Daphne Major. El pinzón medio molido se alimenta de semillas. Las aves han heredado variaciones en la forma del pico, con algunos individuos que tienen pico ancho y profundo y otros con pico más delgado. Las aves de pico grande se alimentan de manera más eficiente con semillas grandes y duras, mientras que las aves de pico más pequeño se alimentan de manera más eficiente con semillas pequeñas y blandas. Durante 1977, un periodo de sequía alteró la vegetación de la isla. Después de este periodo, el número de semillas disminuyó drásticamente: la disminución de las semillas pequeñas y blandas fue mayor que la disminución de las semillas grandes y duras. Las aves de pico grande pudieron sobrevivir mejor que las aves de pico pequeño al año siguiente. Al año siguiente a la sequía cuando los Grants midieron tamaños de pico en la población muy reducida, encontraron que el tamaño promedio de los billetes era mayor (Figura\(\PageIndex{3}\)). Esta fue evidencia clara para la selección natural (diferencias en la supervivencia) del tamaño del pico causada por la disponibilidad de semillas. Los Grants habían estudiado la herencia de tamaños de pico y sabían que las aves supervivientes de pico grande tenderían a producir crías con pico más grande, por lo que la selección conduciría a la evolución del tamaño del pico. Estudios posteriores de los Grants han demostrado la selección y evolución del tamaño del pico en esta especie en respuesta a las condiciones cambiantes en la isla. La evolución se ha dado tanto a billetes más grandes, como en este caso, como a billetes más pequeños cuando las semillas grandes se volvieron raras.
Variación y Adaptación
La selección natural sólo puede realizarse si hay variación, o diferencias, entre los individuos de una población. Es importante destacar que estas diferencias deben tener alguna base genética; de lo contrario, la selección no conducirá a cambios en la próxima generación. Esto es crítico porque la variación entre los individuos puede ser causada por razones no genéticas, como que un individuo sea más alto debido a una mejor nutrición que a diferentes genes.
La diversidad genética en una población proviene de dos fuentes principales: la mutación y la reproducción sexual. La mutación, un cambio en el ADN, es la fuente definitiva de nuevos alelos o nuevas variaciones genéticas en cualquier población. Un individuo que tiene un gen mutado podría tener un rasgo diferente al de otros individuos en la población. No obstante, no siempre es así. Una mutación puede tener uno de tres resultados en la apariencia (o fenotipo) de los organismos:
- Una mutación puede afectar el fenotipo del organismo de una manera que le da forma reducida, menor probabilidad de supervivencia, lo que resulta en menos descendencia.
- Una mutación puede producir un fenotipo con un efecto beneficioso sobre la condición física.
- Muchas mutaciones, llamadas mutaciones neutras, no tendrán ningún efecto sobre la aptitud física.
Las mutaciones también pueden tener toda una gama de tamaños de efecto sobre la aptitud del organismo que las expresa en su fenotipo, desde un pequeño efecto hasta un gran efecto. La reproducción sexual y el cruce en la meiosis también conducen a la diversidad genética: cuando dos padres se reproducen, se ensamblan combinaciones únicas de alelos para producir genotipos únicos y, por lo tanto, fenotipos en cada una de las crías.
Un rasgo heredable que ayuda a la supervivencia y reproducción de un organismo en su ambiente actual se llama adaptación. Una adaptación es una “coincidencia” del organismo con el medio ambiente. La adaptación a un ambiente se produce cuando se produce un cambio en el rango de variación genética a lo largo del tiempo que aumenta o mantiene la correspondencia de la población con su entorno. Las variaciones en los picos de pinzón cambiaron de generación en generación proporcionando adaptación a la disponibilidad de alimentos.
Que un rasgo sea favorable o no depende del entorno en ese momento. Los mismos rasgos no siempre tienen el mismo beneficio relativo o desventaja porque las condiciones ambientales pueden cambiar. Por ejemplo, los pinzones con billetes grandes se beneficiaron en un clima, mientras que los billetes pequeños fueron una desventaja; en un clima diferente, la relación se invirtió.
Patrones de Evolución
La evolución de las especies ha dado lugar a una enorme variación en forma y función. Cuando dos especies evolucionan en diferentes direcciones desde un punto común, se llama evolución divergente. Tal evolución divergente se puede observar en las formas de los órganos reproductivos de las plantas con flores, que comparten las mismas anatomías básicas; sin embargo, pueden verse muy diferentes como resultado de la selección en diferentes ambientes físicos, y la adaptación a diferentes tipos de polinizadores (Figura\(\PageIndex{4}\)).

En otros casos, fenotipos similares evolucionan independientemente en especies distantes relacionadas. Por ejemplo, el vuelo ha evolucionado tanto en murciélagos como en insectos, y ambos tienen estructuras a las que nos referimos como alas, que son adaptaciones al vuelo. Las alas de murciélagos e insectos, sin embargo, evolucionaron a partir de estructuras originales muy diferentes. Cuando estructuras similares surgen a través de la evolución independientemente en diferentes especies se llama evolución convergente. Las alas de murciélagos e insectos se denominan estructuras análogas; son similares en función y apariencia, pero no comparten un origen en un ancestro común. En cambio evolucionaron independientemente en los dos linajes. Las alas de un colibrí y un avestruz son estructuras homólogas, lo que significa que comparten similitudes (a pesar de sus diferencias resultantes de divergencia evolutiva). Las alas de colibríes y avestruces no evolucionaron independientemente en el linaje de colibríes y avestruces, descendieron de un ancestro común con alas.
La síntesis moderna
Los mecanismos de herencia, la genética, no se entendieron en el momento en que Darwin y Wallace estaban desarrollando su idea de selección natural. Esta falta de comprensión fue un obstáculo para comprender muchos aspectos de la evolución. De hecho, la herencia de mezcla era la teoría genética predominante (e incorrecta) de la época, lo que dificultaba la comprensión de cómo podría operar la selección natural. Darwin y Wallace desconocían el trabajo genético del monje austriaco Gregor Mendel, que se publicó en 1866, poco después de la publicación de Sobre el origen de las especies. La obra de Mendel fue redescubierta a principios del siglo XX, momento en el que los genetistas llegaban rápidamente a comprender los fundamentos de la herencia. Inicialmente, la naturaleza particulada recién descubierta de los genes dificultaba que los biólogos entendieran cómo podría ocurrir la evolución gradual. Pero en las próximas décadas la genética y la evolución se integraron en lo que se conoció como la síntesis moderna, la comprensión coherente de la relación entre la selección natural y la genética que tomó forma en la década de 1940 y que generalmente se acepta hoy en día. En suma, la síntesis moderna describe cómo las presiones evolutivas, como la selección natural, pueden afectar la composición genética de una población y, a su vez, cómo esto puede resultar en la evolución gradual de poblaciones y especies. La teoría también conecta este cambio gradual de una población a lo largo del tiempo, llamado microevolución, con los procesos que dieron lugar a nuevas especies y grupos taxonómicos superiores con caracteres ampliamente divergentes, llamados macroevolución.
Genética de Poblaciones
Recordemos que un gen para un personaje en particular puede tener varias variantes, o alelos, que codifican para diferentes rasgos asociados a ese carácter. Por ejemplo, en el sistema de tipo sanguíneo ABO en humanos, tres alelos determinan la proteína de tipo sanguíneo particular en la superficie de los glóbulos rojos. Cada individuo en una población de organismos diploides sólo puede portar dos alelos para un gen en particular, pero más de dos pueden estar presentes en los individuos que conforman la población. Mendel siguió alelos ya que fueron heredados de padres a hijos. A principios del siglo XX, los biólogos comenzaron a estudiar qué sucede con todos los alelos en una población en un campo de estudio conocido como genética de poblaciones.
Hasta ahora, hemos definido la evolución como un cambio en las características de una población de organismos, pero detrás de ese cambio fenotípico está el cambio genético. En términos genéticos poblacionales, la evolución se define como un cambio en la frecuencia de un alelo en una población. Usando el sistema ABO como ejemplo, la frecuencia de uno de los alelos, I A, es el número de copias de ese alelo dividido por todas las copias del gen ABO en la población. Por ejemplo, un estudio en Jordania encontró que una frecuencia de I A era 26.1 por ciento. 2 Los alelos I B, I 0 conformaron 13.4 por ciento y 60.5 por ciento de los alelos respectivamente, y todas las frecuencias suman 100 por ciento. Un cambio en esta frecuencia a lo largo del tiempo constituiría evolución en la población.
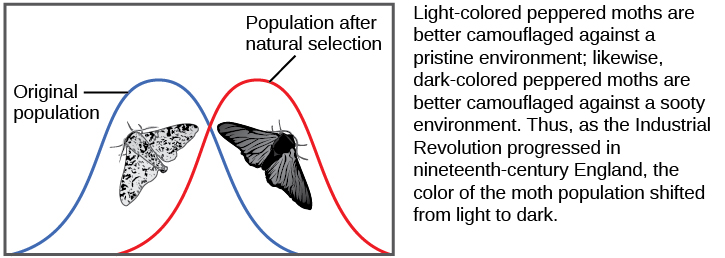
Hay varias maneras en que las frecuencias alélicas de una población pueden cambiar. Una de esas formas es la selección natural. Si un alelo dado confiere un fenotipo que permite que un individuo tenga más descendencia que sobreviva y se reproduzca, ese alelo, en virtud de ser heredado por esas crías, estará en mayor frecuencia en la próxima generación. Dado que las frecuencias alélicas siempre suman el 100 por ciento, un incremento en la frecuencia de un alelo siempre significa una disminución correspondiente en uno o más de los otros alelos. Los alelos altamente beneficiosos pueden, a lo largo de muy pocas generaciones, llegar a ser “fijos” de esta manera, lo que significa que cada individuo de la población portará el alelo. Del mismo modo, los alelos perjudiciales pueden eliminarse rápidamente del acervo genético, la suma de todos los alelos en una población. Parte del estudio de la genética poblacional es rastrear cómo las fuerzas selectivas cambian las frecuencias alélicas en una población a lo largo del tiempo, lo que puede dar a los científicos pistas sobre las fuerzas selectivas que pueden estar operando en una población determinada. Los estudios de cambios en la coloración de las alas en la polilla moteada de blanco moteado a oscuro en respuesta a troncos de árboles cubiertos de hollín y luego de regreso al blanco moteado cuando las fábricas dejaron de producir tanto hollín es un ejemplo clásico de estudio de la evolución en poblaciones naturales (Figura\(\PageIndex{5}\)).
A principios del siglo XX, el matemático inglés Godfrey Hardy y el médico alemán Wilhelm Weinberg proporcionaron de forma independiente una explicación para un concepto algo contradictorio. La explicación original de Hardy fue en respuesta a un malentendido de por qué un alelo “dominante”, uno que enmascara un alelo recesivo, no debería aumentar de frecuencia en una población hasta que eliminó todos los demás alelos. La pregunta resultó de una confusión común sobre lo que significa “dominante”, pero obligó a Hardy, quien ni siquiera era biólogo, a señalar que si no hay factores que afecten una frecuencia alélica esas frecuencias permanecerán constantes de una generación a la siguiente. Este principio se conoce ahora como el equilibrio Hardy-Weinberg. La teoría establece que las frecuencias de alelos y genotipos de una población son inherentemente estables, a menos que algún tipo de fuerza evolutiva esté actuando sobre la población, la población portaría los mismos alelos en las mismas proporciones generación tras generación. Los individuos, en su conjunto, se verían esencialmente iguales y esto no estaría relacionado con si los alelos eran dominantes o recesivos. Las cuatro fuerzas evolutivas más importantes, que perturbarán el equilibrio, son la selección natural, la mutación, la deriva genética y la migración dentro o fuera de una población. Un quinto factor, el apareamiento no aleatorio, también interrumpirá el equilibrio de Hardy-Weinberg pero solo desplazando las frecuencias de genotipos, no las frecuencias alélicas. En el apareamiento no aleatorio, los individuos son más propensos a aparearse con individuos similares (o a diferencia de individuos) en lugar de al azar. Dado que el apareamiento no aleatorio no cambia las frecuencias alélicas, no causa evolución directamente. Se ha descrito la selección natural. La mutación crea un alelo de otro y cambia la frecuencia de un alelo en una cantidad pequeña pero continua en cada generación. Cada alelo es generado por una tasa de mutación baja y constante que aumentará lentamente la frecuencia del alelo en una población si ninguna otra fuerza actúa sobre el alelo. Si la selección natural actúa contra el alelo, se eliminará de la población a una tasa baja conduciendo a una frecuencia que resulta de un equilibrio entre selección y mutación. Esta es una de las razones por las que las enfermedades genéticas permanecen en la población humana a frecuencias muy bajas. Si el alelo se ve favorecido por la selección, aumentará en frecuencia. La deriva genética provoca cambios aleatorios en las frecuencias alélicas cuando las poblaciones son pequeñas. La deriva genética a menudo puede ser importante en la evolución, como se discute en la siguiente sección. Finalmente, si dos poblaciones de una especie tienen diferentes frecuencias alélicas, la migración de individuos entre ellas provocará cambios de frecuencia en ambas poblaciones. Como sucede, no hay población en la que uno o más de estos procesos no estén operando, por lo que las poblaciones siempre están evolucionando, y nunca se observará exactamente el equilibrio Hardy-Weinberg. Sin embargo, el principio Hardy-Weinberg da a los científicos una expectativa de referencia para las frecuencias alélicas en una población no evolutiva con la que pueden comparar poblaciones en evolución y así inferir qué fuerzas evolutivas podrían estar en juego. La población está evolucionando si las frecuencias de alelos o genotipos se desvían del valor esperado del principio Hardy-Weinberg.
Darwin identificó un caso especial de selección natural al que llamó selección sexual. La selección sexual afecta la capacidad de un individuo para aparearse y así producir descendencia, y conduce a la evolución de rasgos dramáticos que a menudo parecen desadaptativos en términos de supervivencia pero persisten porque dan a sus dueños un mayor éxito reproductivo. La selección sexual ocurre de dos maneras: a través de la competencia masculina-masculina por parejas y a través de la selección femenina de parejas. La competencia hombre-hombre toma la forma de conflictos entre hombres, que a menudo se ritualizan, pero también pueden representar amenazas significativas para la supervivencia de un hombre. En ocasiones la competencia es por territorio, con las hembras más propensas a aparearse con machos con territorios de mayor calidad. La elección femenina ocurre cuando las hembras eligen a un macho en función de un rasgo particular, como los colores de las plumas, la realización de una danza de apareamiento o la construcción de una estructura elaborada. En algunos casos, la competencia macho-macho y la elección femenina se combinan en el proceso de apareamiento. En cada uno de estos casos, los rasgos seleccionados para, como la capacidad de combate o el color y la longitud de las plumas, se potencian en los machos. En general, se piensa que la selección sexual puede proceder a un punto en el que la selección natural contra la mejora adicional de un personaje impide su evolución posterior porque impacta negativamente en la capacidad del macho para sobrevivir. Por ejemplo, plumas coloridas o una elaborada exhibición hacen que el macho sea más obvio para los depredadores.
Resumen
La evolución por selección natural surge de tres condiciones: los individuos dentro de una especie varían, algunas de esas variaciones son heredables y los organismos tienen más descendencia de la que los recursos pueden soportar. La consecuencia es que los individuos con variaciones relativamente ventajosas tendrán más probabilidades de sobrevivir y tendrán tasas reproductivas más altas que aquellos individuos con rasgos diferentes. Los rasgos ventajosos se transmitirán a la descendencia en mayor proporción. De esta manera, el rasgo tendrá una mayor representación en las generaciones siguientes y posteriores, lo que conducirá a un cambio genético en la población.
La síntesis moderna de la teoría evolutiva surgió de la reconciliación de los pensamientos de Darwin, Wallace y Mendel sobre la evolución y la herencia. La genética poblacional es un marco teórico para describir el cambio evolutivo en las poblaciones a través del cambio en las frecuencias alélicas. La genética poblacional define la evolución como un cambio en la frecuencia de los alelos durante generaciones. En ausencia de fuerzas evolutivas, las frecuencias alélicas no cambiarán en una población; esto se conoce como principio de equilibrio Hardy-Weinberg. Sin embargo, en todas las poblaciones, la mutación, la selección natural, la deriva genética y la migración actúan para cambiar las frecuencias alélicas.
Notas al pie
- 1 Charles Darwin, Revista de Investigaciones sobre la Historia Natural y Geología de los Países Visitados durante el Viaje de H.M.S. Beagle Alrededor del Mundo, bajo el Mando del Capt. Fitz Roy, R.N, 2da. ed. (Londres: John Murray, 1860), http://www.archive.org/details/journalofresea00darw.
- 2 Sahar S. Hanania, Dhia S. Hassawi, y Nidal M. Irshaid, “Frecuencia alélica y genotipos moleculares del sistema de grupos sanguíneos ABO en una población jordana”, Journal of Medical Sciences 7 (2007): 51-58, doi:10.3923/jms.2007.51.58
Glosario
- adaptación
- un rasgo o comportamiento heredable en un organismo que ayuda a su supervivencia en su entorno actual
- estructura análoga
- una estructura que es similar debido a la evolución en respuesta a presiones de selección similares que dan como resultado una evolución convergente, no similar por el descenso de un ancestro común
- evolución convergente
- una evolución que da como resultado formas similares en diferentes especies
- evolución divergente
- una evolución que da como resultado diferentes formas en dos especies con un ancestro común
- acervo genético
- todos los alelos portados por todos los individuos de la población
- deriva genética
- el efecto del azar en el acervo genético de una población
- estructura homóloga
- una estructura que es similar debido a la descendencia de un ancestro común
- herencia de características adquiridas
- una frase que describe el mecanismo de evolución propuesto por Lamarck en el que los rasgos adquiridos por los individuos a través del uso o desuso podrían transmitirse a su descendencia conduciendo así a un cambio evolutivo en la población
- macroevolución
- una escala más amplia de cambios evolutivos vistos a lo largo del tiempo paleontológico
- microevolución
- los cambios en la estructura genética de una población (es decir, frecuencia alélica)
- migración
- el movimiento de individuos de una población a una nueva ubicación; en genética poblacional se refiere al movimiento de individuos y sus alelos de una población a otra, potencialmente cambiando las frecuencias alélicas tanto en la población antigua como en la nueva
- síntesis moderna
- el paradigma evolutivo global que tomó forma en la década de 1940 y es generalmente aceptado hoy
- selección natural
- la mayor supervivencia relativa y reproducción de individuos en una población que tiene rasgos heredables favorables, lo que lleva a un cambio evolutivo
- genética poblacional
- el estudio de cómo las fuerzas selectivas cambian las frecuencias alélicas en una población a lo largo del tiempo
- variación
- la variedad de alelos en una población