
17.2: Inmunidad innata
- Page ID
- 53514

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)El sistema inmune vertebrado, incluido el humano, es un complejo sistema multicapa para defenderse de amenazas externas e internas a la integridad del cuerpo. El sistema se puede dividir en dos tipos de sistemas de defensa: el sistema inmune innato, que es inespecífico hacia un tipo particular de patógeno, y el sistema inmune adaptativo, que es específico (Figura\(\PageIndex{1}\)). La inmunidad innata no es causada por una infección o vacunación y depende inicialmente de barreras físicas y químicas que funcionan sobre todos los patógenos, a veces llamados la primera línea de defensa. La segunda línea de defensa del sistema innato incluye señales químicas que producen respuestas de inflamación y fiebre así como la movilización de células protectoras y otras defensas químicas. El sistema inmune adaptativo monta una respuesta altamente específica a sustancias y organismos que no pertenecen al organismo. El sistema adaptativo tarda más en responder y tiene un sistema de memoria que le permite responder con mayor intensidad en caso de que el cuerpo reencuentre un patógeno incluso años después.

Barreras externas y químicas
El cuerpo tiene barreras físicas significativas para los posibles patógenos. La piel contiene la proteína queratina, que resiste la entrada física en las células. Otras superficies corporales, particularmente las asociadas con las aberturas corporales, están protegidas por las membranas mucosas. El moco pegajoso proporciona una trampa física para los patógenos, impidiendo su movimiento más profundo en el cuerpo. Las aberturas del cuerpo, como la nariz y las orejas, están protegidas por pelos que atrapan patógenos, y las membranas mucosas del tracto respiratorio superior tienen cilios que mueven constantemente los patógenos atrapados en la capa mucosa hasta la boca.
La piel y las membranas mucosas también crean un ambiente químico que es hostil a muchos microorganismos. La superficie de la piel es ácida, lo que impide el crecimiento bacteriano. La saliva, el moco y las lágrimas del ojo contienen una enzima que descompone las paredes celulares bacterianas. Las secreciones estomacales crean un ambiente altamente ácido, que mata a muchos patógenos que ingresan al sistema digestivo.
Finalmente, la superficie del cuerpo y el sistema digestivo inferior tienen una comunidad de microorganismos como bacterias, arqueas y hongos que coexisten sin dañar el cuerpo. Existe evidencia de que estos organismos son altamente beneficiosos para su hospedador, combatiendo a los organismos causantes de enfermedades y superándolos por los recursos nutricionales proporcionados por el organismo huésped. A pesar de estas defensas, los patógenos pueden ingresar al organismo a través de abrasiones o pinchazos cutáneos, o por la recolección en superficies mucosas en grandes cantidades que superan las protecciones de moco o cilios.
Defensas internas
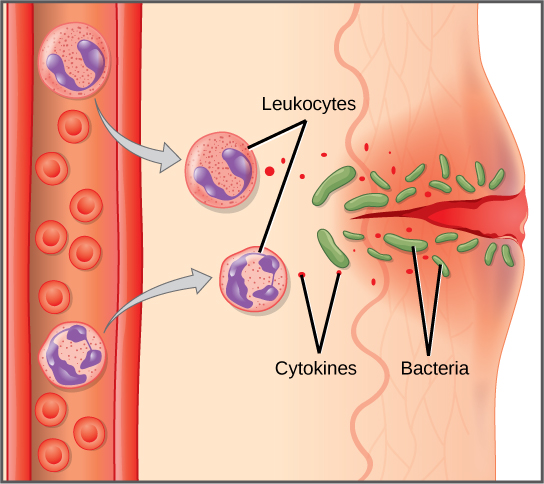
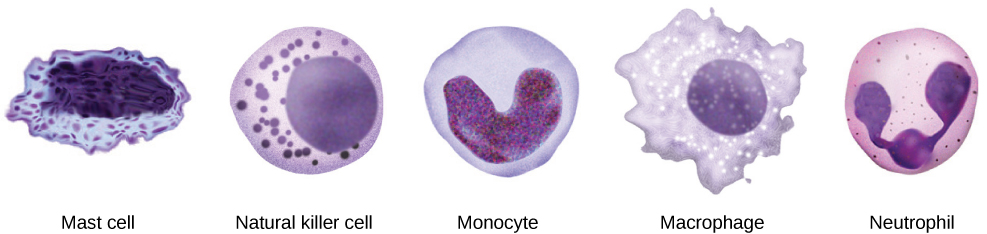
Cuando los patógenos ingresan al cuerpo, el sistema inmunitario innato responde con una variedad de defensas internas. Estos incluyen la respuesta inflamatoria, fagocitosis, células asesinas naturales y el sistema del complemento. Los glóbulos blancos en la sangre y la linfa reconocen a los patógenos como extraños al cuerpo. Un glóbulo blanco es más grande que un glóbulo rojo, está nucleado y, por lo general, es capaz de moverse usando la locomoción ameboidea. Debido a que pueden moverse por su cuenta, los glóbulos blancos pueden dejar la sangre para ir a los tejidos infectados. Por ejemplo, un monocito es un tipo de glóbulo blanco que circula en la sangre y la linfa y se convierte en un macrófago después de que se mueve hacia el tejido infectado. Un macrófago es una célula grande que envuelve partículas extrañas y patógenos. Los mastocitos se producen de la misma manera que los glóbulos blancos, pero a diferencia de los glóbulos blancos circulantes, los mastocitos toman residencia en los tejidos conectivos y especialmente en los tejidos mucosos. Se encargan de liberar químicos en respuesta a lesiones físicas. También juegan un papel en la respuesta alérgica, que se discutirá más adelante en el capítulo.
Cuando un patógeno es reconocido como extraño, se liberan sustancias químicas llamadas citoquinas. Una citocina es un mensajero químico que regula la diferenciación celular (forma y función), la proliferación (producción) y la expresión génica para producir una variedad de respuestas inmunitarias. Aproximadamente 40 tipos de citocinas existen en humanos. Además de ser liberadas de los glóbulos blancos después del reconocimiento de patógenos, las citocinas también son liberadas por las células infectadas y se unen a células no infectadas cercanas, induciendo a esas células a liberar citocinas. Este bucle de retroalimentación positiva da como resultado una explosión de producción de citocinas.
Una clase de citocinas de acción temprana son los interferones, que son liberados por las células infectadas como una advertencia para las células cercanas no infectadas. Un interferón es una proteína pequeña que señala una infección viral a otras células. Los interferones estimulan a las células no infectadas para producir compuestos que interfieren con la replicación viral. Los interferones también activan macrófagos y otras células.
La respuesta inflamatoria y la fagocitosis
Las primeras citocinas que se producen fomentan la inflamación, enrojecimiento localizado, hinchazón, calor y dolor. La inflamación es una respuesta a un trauma físico, como un corte o un golpe, irritación química e infección por patógenos (virus, bacterias u hongos). Las señales químicas que desencadenan una respuesta inflamatoria ingresan al líquido extracelular y hacen que los capilares se dilaten (expandan) y las paredes capilares se vuelvan más permeables o con fugas. El suero y otros compuestos que se escapan de los capilares provocan hinchazón de la zona, lo que a su vez causa dolor. Diversos tipos de glóbulos blancos son atraídos por el área de inflamación. Los tipos de glóbulos blancos que llegan a un sitio inflamado dependen de la naturaleza de la lesión o patógeno infeccioso. Por ejemplo, un neutrófilo es un glóbulo blanco que llega temprano que engulle y digiere patógenos. Los neutrófilos son los glóbulos blancos más abundantes del sistema inmune (Figura\(\PageIndex{2}\)). Los macrófagos siguen a los neutrófilos y toman la función de fagocitosis y están involucrados en la resolución de un sitio inflamado, limpiando restos celulares y patógenos.
Las citocinas también envían retroalimentación a las células del sistema nervioso para provocar los síntomas generales de sentirse enfermo, que incluyen letargo, dolor muscular y náuseas. Las citocinas también aumentan la temperatura corporal central, provocando fiebre. Las elevadas temperaturas de una fiebre inhiben el crecimiento de patógenos y aceleran los procesos de reparación celular. Por estas razones, la supresión de las fiebres debe limitarse a las que son peligrosamente altas.
CONCEPT EN ACCIÓN
Echa un vistazo a este video stop-motion de 23 segundos que muestra un neutrófilo que busca y envuelve esporas de hongos durante un tiempo transcurrido de 79 minutos.
Células asesinas naturales
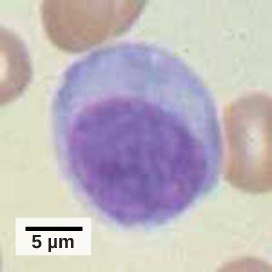
Un linfocito es un glóbulo blanco que contiene un núcleo grande (Figura\(\PageIndex{3}\)). La mayoría de los linfocitos están asociados con la respuesta inmune adaptativa, pero las células infectadas son identificadas y destruidas por las células asesinas naturales, los únicos linfocitos del sistema inmune innato. Una célula asesina natural (NK) es un linfocito que puede matar células infectadas con virus (o células cancerosas). Las células NK identifican infecciones intracelulares, especialmente por virus, por la expresión alterada de moléculas de clase mayor de histocompatibilidad (MHC) I en la superficie de las células infectadas. Las moléculas MHC clase I son proteínas en las superficies de todas las células nucleadas que proporcionan una muestra del entorno interno de la célula en un momento dado. Las células poco saludables, ya sean infectadas o cancerosas, muestran un complemento alterado del MHC clase I en sus superficies celulares.
Después de que la célula NK detecta una célula infectada o tumoral, induce la muerte celular programada, o apoptosis. Luego, las células fagocíticas vienen y digieren los restos celulares que quedan atrás. Las células NK patrullan constantemente el cuerpo y son un mecanismo eficaz para controlar posibles infecciones y prevenir la progresión del cáncer. Los diversos tipos de células inmunes se muestran en la Figura\(\PageIndex{4}\).
Complemento
Una matriz de aproximadamente 20 tipos de proteínas, llamadas sistema del complemento, también se activa por la infección o la actividad de las células del sistema inmune adaptativo y funciona para destruir los patógenos extracelulares. Las células hepáticas y los macrófagos sintetizan formas inactivas de proteínas del complemento continuamente; estas proteínas son abundantes en el suero sanguíneo y son capaces de responder inmediatamente a microorganismos infectantes. El sistema del complemento se llama así porque es complementario al sistema inmune innato y adaptativo. Las proteínas del complemento se unen a las superficies de los microorganismos y son particularmente atraídas por patógenos que ya están etiquetados por el sistema inmune adaptativo. Este “etiquetado” implica la unión de proteínas específicas llamadas anticuerpos (discutidas en detalle más adelante) al patógeno. Cuando se unen, los anticuerpos cambian de forma proporcionando un sitio de unión para una de las proteínas del complemento. Después de que las primeras proteínas del complemento se unen, sigue una cascada de unión en una secuencia específica de proteínas en la que el patógeno se recubre rápidamente con proteínas del complemento.
Las proteínas del complemento realizan varias funciones, una de las cuales es servir como marcador para indicar la presencia de un patógeno a las células fagocíticas y potenciar el engullido. Ciertas proteínas del complemento pueden combinarse para abrir los poros en las membranas celulares microbianas y causar lisis de las células.
Resumen
El sistema inmune innato consiste primero en barreras físicas y químicas a la infección, incluyendo la piel y las membranas mucosas y sus secreciones, superficies ciliadas y pelos corporales. La segunda línea de defensa es un sistema de defensa interna diseñado para contrarrestar las amenazas patógenas que pasan por alto las barreras físicas y químicas del cuerpo. Mediante una combinación de respuestas celulares y moleculares, el sistema inmune innato identifica la naturaleza de un patógeno y responde con inflamación, fagocitosis, liberación de citocinas, destrucción por células NK o el sistema del complemento.
Glosario
- sistema de complemento
- una matriz de aproximadamente 20 proteínas solubles del sistema inmune innato que mejoran la fagocitosis, perforan los patógenos y reclutan linfocitos
- citocina
- un mensajero químico que regula la diferenciación celular, la proliferación y la expresión génica para efectuar respuestas inmunitarias
- inflamación
- el enrojecimiento localizado, hinchazón, calor y dolor que resulta del movimiento de leucocitos a través de capilares abiertos a un sitio de infección
- inmunidad innata
- una inmunidad que ocurre naturalmente por factores genéticos o fisiología, y no es causada por infección o vacunación
- interferón
- una citocina que inhibe la replicación viral
- linfocito
- un tipo de glóbulo blanco que incluye células asesinas naturales del sistema inmune innato y células B y T del sistema inmune adaptativo
- macrófagos
- una célula fagocítica grande que envuelve partículas extrañas y patógenos
- clase mayor de histocompatibilidad (MHC) I
- un grupo de proteínas que se encuentran en la superficie de todas las células nucleadas que señalan a las células inmunitarias si la célula es normal o está infectada o cancerosa; también proporciona los sitios apropiados en los que los antígenos pueden cargarse para su reconocimiento por los linfocitos
- mastocitos
- un leucocito que produce moléculas inflamatorias, como la histamina, en respuesta a grandes patógenos
- monocito
- un tipo de glóbulo blanco que circula en la sangre y la linfa y se diferencia en un macrófago después de que se mueve hacia el tejido infectado
- célula asesina natural (NK)
- un linfocito que puede destruir células infectadas con virus o células tumorales
- neutrófilos
- un leucocito fagocítico que engulle y digiere patógenos
- glóbulo blanco
- una célula nucleada que se encuentra en la sangre que forma parte del sistema inmune; también llamada leucocitos