
7.2: Tipos de receptores
- Page ID
- 58430
Una célula dentro de un organismo multicelular puede necesitar señalizar a otras células que se encuentran a varias distancias de la célula original (Figura\(\PageIndex{1}\)). No todas las células se ven afectadas por las mismas señales. Se utilizan diferentes tipos de señalización para diferentes propósitos.
Los receptores son moléculas proteicas dentro de la célula diana o en su superficie que reciben una señal química. Las señales químicas son liberadas por las células de señalización en forma de moléculas pequeñas, generalmente volátiles o solubles llamadas ligandos. Un ligando es una molécula que se une a otra molécula específica, en algunos casos, entregando una señal en el proceso. Por lo tanto, los ligandos pueden pensarse como moléculas de señalización. Los ligandos y receptores existen en varias variedades; sin embargo, un ligando específico tendrá un receptor específico que típicamente se une solo a ese ligando.
Receptores internos
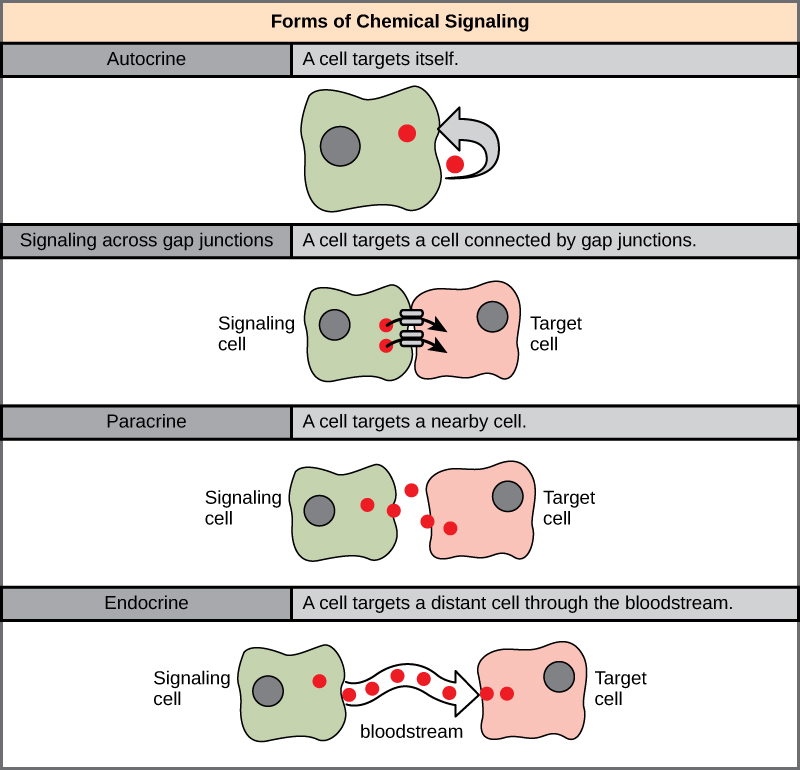
Los receptores internos, también conocidos como receptores intracelulares o citoplásmicos, se encuentran en el citoplasma de la célula y responden a moléculas de ligando hidrófobas que son capaces de viajar a través de la membrana plasmática. Una vez dentro de la célula, muchas de estas moléculas se unen a proteínas que actúan como reguladores de la síntesis de ARNm. Recordemos que el ARNm transporta información genética desde el ADN en el núcleo de una célula hasta el ribosoma, donde se ensambla la proteína. Cuando el ligando se une al receptor interno, se desencadena un cambio de forma que expone un sitio de unión a ADN en la proteína receptora. El complejo ligando-receptor se mueve hacia el núcleo, luego se une a regiones específicas del ADN y promueve la producción de ARNm a partir de genes específicos (Figura\(\PageIndex{2}\)). Los receptores internos pueden influir directamente en la expresión génica (la cantidad de una proteína específica que se produce a partir de un gen) sin tener que pasar la señal a otros receptores o mensajeros.
Receptores de superficie celular
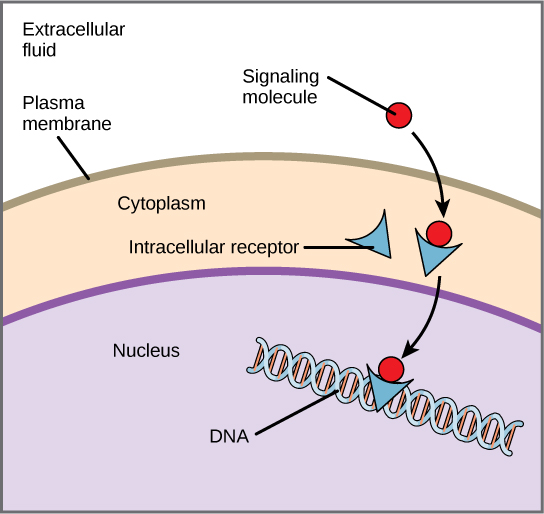
Los receptores de superficie celular, también conocidos como receptores transmembrana, son proteínas que se encuentran unidas a la membrana celular. Estos receptores se unen a moléculas de ligando externas (ligandos que no viajan a través de la membrana celular). Este tipo de receptor abarca la membrana plasmática y realiza la transducción de señales, en la que una señal extracelular se convierte en una señal intercelular. Los ligandos que interactúan con los receptores de la superficie celular no tienen que ingresar a la célula a la que afectan. Los receptores de superficie celular también se denominan proteínas o marcadores específicos de células porque son específicos de tipos celulares individuales.
Cada receptor de superficie celular tiene tres componentes principales: un dominio externo de unión a ligando, una región hidrofóbica que abarca la membrana y un dominio intracelular dentro de la célula. El tamaño y extensión de cada uno de estos dominios varían ampliamente, dependiendo del tipo de receptor.
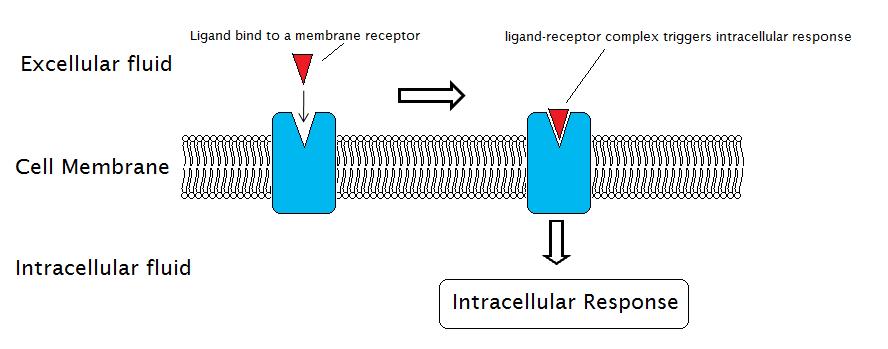
Receptores enlazados a canales iónicos
Los receptores unidos a canales iónicos se unen a un ligando y abren un canal a través de la membrana que permite el paso de iones específicos. Para formar un canal, este tipo de receptor de superficie celular tiene una extensa región que abarca la membrana. Cuando un ligando se une a la región extracelular del canal, hay un cambio conformacional en la estructura de las proteínas que permite el paso de iones como sodio, calcio, magnesio e hidrógeno (Figura\(\PageIndex{4}\)).
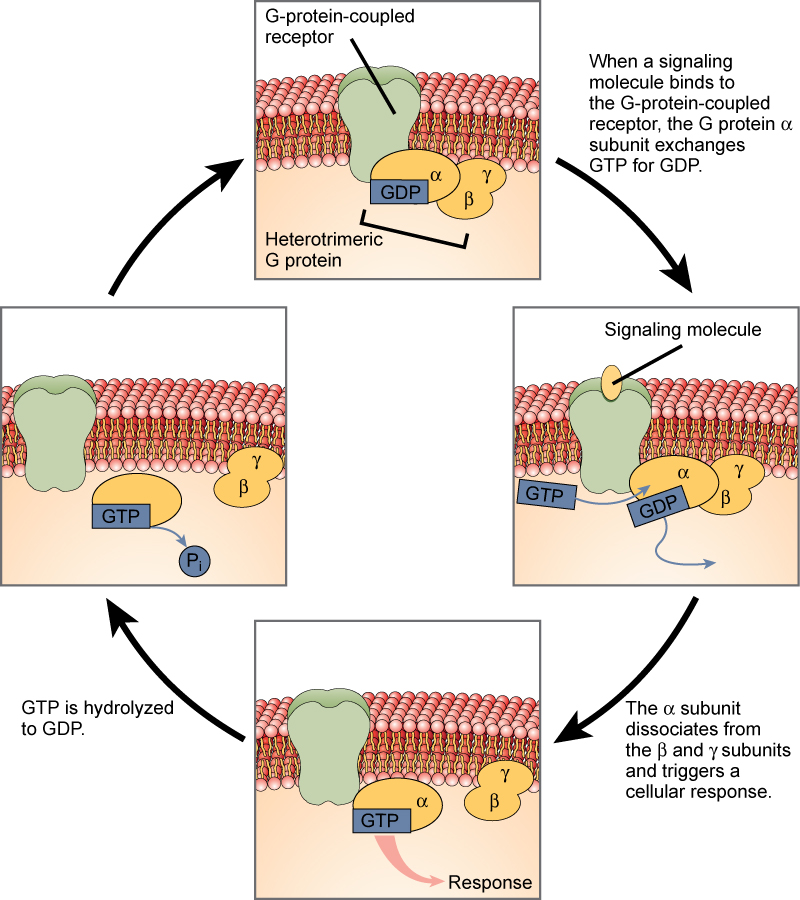
Receptores acoplados a proteína G
Los receptores acoplados a proteína G se unen a un ligando y activan una proteína de membrana llamada proteína G. La proteína G activada interactúa entonces con un canal iónico o una enzima en la membrana (Figura\(\PageIndex{5}\)). Antes de que el ligando se una, la proteína G inactiva puede unirse a un sitio en un receptor específico. Una vez que la proteína G se une al receptor, la proteína G cambia de forma, se activa y se divide en dos subunidades diferentes. Una o ambas de estas subunidades pueden ser capaces de activar otras proteínas como resultado.
Receptores ligados a enzimas
Los receptores ligados a enzimas son receptores de superficie celular con dominios intracelulares que están asociados con una enzima. En algunos casos, el dominio intracelular del receptor en sí es una enzima. Otros receptores ligados a enzimas tienen un pequeño dominio intracelular que interactúa directamente con una enzima. Cuando un ligando se une al dominio extracelular, se transfiere una señal a través de la membrana, activando la enzima. La activación de la enzima desencadena una cadena de eventos dentro de la célula que eventualmente conduce a una respuesta.
Cómo los virus reconocen a un anfitrión
A diferencia de las células vivas, muchos virus no tienen una membrana plasmática ni ninguna de las estructuras necesarias para sostener la vida. Algunos virus se componen simplemente de una cubierta proteica inerte que contiene ADN o ARN. Para reproducirse, los virus deben invadir una célula viva, que sirve como huésped, para luego hacerse cargo del aparato celular hospedador. Pero, ¿cómo reconoce un virus a su huésped?
Los virus a menudo se unen a los receptores de la superficie celular en la célula huésped. Por ejemplo, el virus que causa la influenza humana (gripe) se une específicamente a receptores en las membranas de las células del sistema respiratorio. Las diferencias químicas en los receptores de la superficie celular entre los huéspedes significan que un virus que infecta a una especie específica (por ejemplo, humanos) no puede infectar a otra especie (por ejemplo, pollos).
Sin embargo, los virus tienen cantidades muy pequeñas de ADN o ARN en comparación con los humanos y, como resultado, la reproducción viral puede ocurrir rápidamente. La reproducción viral produce invariablemente errores que pueden conducir a cambios en los virus recién producidos; estos cambios significan que las proteínas virales que interactúan con los receptores de la superficie celular pueden evolucionar de tal manera que pueden unirse a receptores en un nuevo huésped. Tales cambios ocurren aleatoriamente y con bastante frecuencia en el ciclo reproductivo de un virus, pero los cambios solo importan si un virus con nuevas propiedades de unión entra en contacto con un hospedador adecuado. En el caso de la influenza, esta situación puede ocurrir en entornos donde animales y personas están en contacto cercano, como granjas avícolas y porcinas (Sigalov, 2010). Una vez que un virus salta a un nuevo host, puede propagarse rápidamente. Los científicos observan de cerca los virus que aparecen recientemente (llamados virus emergentes) con la esperanza de que dicho monitoreo pueda reducir la probabilidad de epidemias virales globales.
Referencias
Texto adaptado de: OpenStax, Biología. OpenStax CNX. 13 de octubre de 2017. https://cnx.org/contents/GFy_h8cu@10...llul#footnote1
A. B. Sigalov, La Escuela de la Naturaleza. IV. Aprendiendo de los virus, Self/Nonself 1, núm. 4 (2010): 282-298. Y. Cao, X. Koh, L. Dong, X. Du, A. Wu, X. Ding, H. Deng, Y. Shu, J. Chen, T. Jiang, Estimación rápida de la actividad de unión de la hemaglutinina del virus de la influenza a receptores humanos y aviares, PLoS One 6, núm. 4 (2011): e18664.