
7.5: Respuesta a la señal
- Page ID
- 58446
Los ligandos que pueden ingresar a la célula y unirse a receptores internos son capaces de afectar directamente el ADN de la célula y la maquinaria productora de proteínas. Los ligandos que no pueden entrar en la célula se unen a receptores en la membrana plasmática y utilizan vías de transducción de señales para producir una variedad de efectos. Los resultados de las vías de señalización son extremadamente variados y dependen del tipo de célula involucrada, así como de las condiciones externas e internas. A continuación se describe una pequeña muestra de respuestas.
Expresión Génica
Algunas vías de transducción de señales regulan la producción de ARNm. Otros regulan la síntesis de proteínas a partir de ARNm por ribosomas. Por lo general, estas vías aumentan la expresión génica de modo que se produce más proteína específica a partir de un gen, pero algunas vías disminuyen la expresión génica.
Un ejemplo de una proteína que regula la traducción en el núcleo es la MAP quinasa ERK. La ERK se activa en una cascada de fosforilación cuando el factor de crecimiento epidérmico (EGF) se une al receptor de EGF. Tras la fosforilación, ERK ingresa al núcleo y activa una proteína quinasa que, a su vez, regula la traducción de proteínas (Figura\(\PageIndex{1}\)).
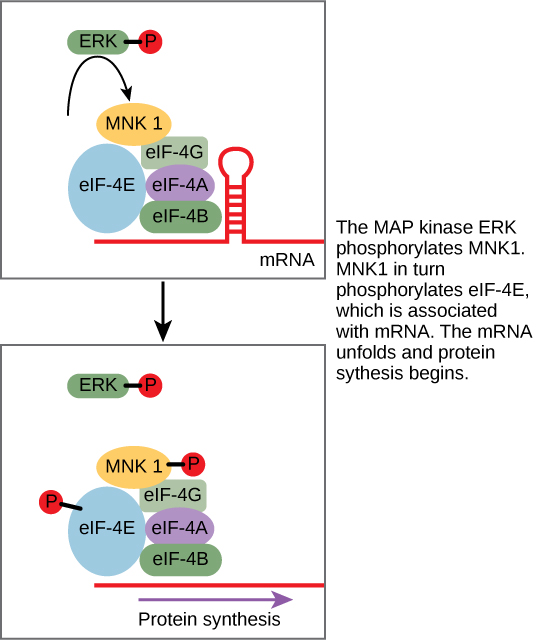
Aumento del Metabolismo Celular
El resultado de otra vía de señalización afecta a las células musculares. La activación de los receptores β-adrenérgicos en las células musculares por la adrenalina conduce a un aumento del AMP cíclico (AMPc) dentro de la célula. También conocida como epinefrina, la adrenalina es una hormona (producida por la glándula suprarrenal adherida al riñón) que prepara al cuerpo para emergencias a corto plazo. El AMP cíclico activa la PKA (proteína quinasa A), que a su vez fosforila dos enzimas. La primera enzima promueve la degradación del glucógeno activando la glucógeno fosforilasa quinasa intermedia (GPK) que a su vez activa la glucógeno fosforilasa (GP) que cataboliza el glucógeno en glucosa. (Recuerda que tu cuerpo convierte el exceso de glucosa en glucógeno para almacenamiento a corto plazo. Cuando se necesita energía, el glucógeno se reconvierte rápidamente en glucosa). La fosforilación de la segunda enzima, la glucógeno sintasa (GS), inhibe su capacidad para formar glucógeno a partir de la glucosa. De esta manera, una célula muscular obtiene un charco listo de glucosa activando su formación a través de la degradación del glucógeno e inhibiendo el uso de glucosa para formar glucógeno, evitando así un ciclo inútil de degradación y síntesis del glucógeno. La glucosa está entonces disponible para su uso por la célula muscular en respuesta a una repentina oleada de adrenalina, el reflejo de “lucha o huida”.
Crecimiento Celular
Las vías de señalización celular también juegan un papel importante en la división celular. Las células normalmente no se dividen a menos que sean estimuladas por señales de otras células. Los ligandos que promueven el crecimiento celular se denominan factores de crecimiento. La mayoría de los factores de crecimiento se unen a receptores de superficie celular que están vinculados a tirosina quinasas. Estos receptores de superficie celular se denominan receptores tirosina quinasas (RTK). La activación de RTK inicia una vía de señalización que incluye una proteína G llamada RAS, que activa la ruta MAP quinasa descrita anteriormente. La enzima MAP quinasa estimula entonces la expresión de proteínas que interactúan con otros componentes celulares para iniciar la división celular.
Muerte Celular
Cuando una célula está dañada, es superflua o potencialmente peligrosa para un organismo, una célula puede iniciar un mecanismo para desencadenar la muerte celular programada, o apoptosis. La apoptosis permite que una célula muera de manera controlada que impide la liberación de moléculas potencialmente dañinas desde el interior de la célula. Existen muchos puntos de control internos que monitorean la salud de una célula; si se observan anomalías, una célula puede iniciar espontáneamente el proceso de apoptosis. Sin embargo, en algunos casos, como una infección viral o una división celular incontrolada por cáncer, fallan los controles y equilibrios normales de la célula. La señalización externa también puede iniciar la apoptosis. Por ejemplo, la mayoría de las células animales normales tienen receptores que interactúan con la matriz extracelular, una red de glicoproteínas que proporciona soporte estructural a las células de un organismo. La unión de los receptores celulares a la matriz extracelular inicia una cascada de señalización dentro de la célula. Sin embargo, si la célula se aleja de la matriz extracelular, la señalización cesa y la célula sufre apoptosis. Este sistema evita que las células viajen por el cuerpo y proliferen fuera de control, como sucede con las células tumorales que metastatizan.
Otro ejemplo de señalización externa que conduce a la apoptosis ocurre en el desarrollo de células T. Las células T son células inmunes que se unen a macromoléculas y partículas extrañas, y se dirigen a ellas para su destrucción por parte del sistema inmune. Normalmente, las células T no se dirigen a proteínas “propias” (las de su propio organismo), un proceso que puede conducir a enfermedades autoinmunes. Con el fin de desarrollar la capacidad de discriminar entre uno mismo y lo no propio, las células T inmaduras se someten a cribado para determinar si se unen a las llamadas autoproteínas. Si el receptor de células T se une a proteínas propias, la célula inicia la apoptosis para eliminar la célula potencialmente peligrosa.
La apoptosis también es esencial para el desarrollo embriológico normal. En los vertebrados, por ejemplo, las primeras etapas de desarrollo incluyen la formación de tejido similar a una banda entre los dedos de las manos y los pies individuales (Figura\(\PageIndex{2}\)). Durante el transcurso del desarrollo normal, estas células innecesarias deben ser eliminadas, permitiendo que se formen dedos de manos y pies completamente separados. Un mecanismo de señalización celular desencadena la apoptosis, que destruye las células entre los dígitos en desarrollo.
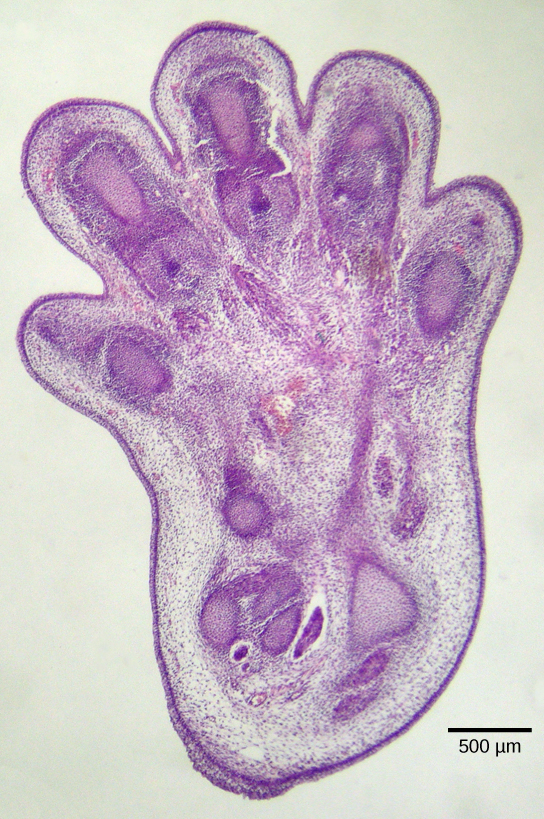
Terminación de la Cascada de Señales
La señalización aberrante que a menudo se observa en las células tumorales es una prueba de que la terminación de una señal en el momento apropiado puede ser tan importante como el inicio de una señal. Un método para detener una señal específica es degradar el ligando o eliminarlo para que ya no pueda acceder a su receptor. Una razón por la que las hormonas hidrofóbicas como el estrógeno y la testosterona desencadenan eventos duraderos es porque se unen a proteínas Estas proteínas permiten que las moléculas insolubles sean solubles en sangre, pero también protegen a las hormonas de la degradación por las enzimas circulantes.
Dentro de la célula, muchas enzimas diferentes revierten las modificaciones celulares que resultan de las cascadas de señalización. Por ejemplo, las fosfatasas son enzimas que eliminan el grupo fosfato unido a las proteínas por las quinasas en un proceso llamado desfosforilación. El AMP cíclico (AMPc) es degradado a AMP por la fosfodiesterasa, y la liberación de depósitos de calcio es revertida por las bombas de Ca 2+ que se localizan en las membranas externa e interna de la célula.
Referencias
OpenStax, Biología. OpenStax CNX. 13 de octubre de 2017. https://cnx.org/contents/GFy_h8cu@10... A la señal