
23.3: Perspectivas sobre el Árbol Filogenético
- Page ID
- 58273
Los conceptos de modelado filogenético cambian constantemente. Es uno de los campos de estudio más dinámicos en toda la biología. En las últimas décadas, nuevas investigaciones han cuestionado las ideas de los científicos sobre cómo se relacionan los organismos. Se han propuesto nuevos modelos de estas relaciones para su consideración por la comunidad científica.
Muchos árboles filogenéticos se han mostrado como modelos de la relación evolutiva entre especies. Los árboles filogenéticos se originaron con Charles Darwin, quien esbozó el primer árbol filogenético en 1837 (Figura\(\PageIndex{1}\) a), que sirvió de patrón para estudios posteriores durante más de un siglo. El concepto de árbol filogenético con un solo tronco que representa a un ancestro común, con las ramas representando la divergencia de especies de este antepasado, encaja bien con la estructura de muchos árboles comunes, como el encino (Figura\(\PageIndex{1}\) b). Sin embargo, la evidencia del moderno análisis de secuencias de ADN y algoritmos informáticos recientemente desarrollados ha causado escepticismo sobre la validez del modelo de árbol estándar en la comunidad científica.

Limitaciones al Modelo Clásico
El pensamiento clásico sobre la evolución procariota, incluido en el modelo de árbol clásico, es que las especies evolucionan clonalmente. Es decir, ellos mismos producen descendencia con mutaciones aleatorias que provocan el descenso a la variedad de especies modernas y extintas conocidas por la ciencia. Esta visión es algo complicada en eucariotas que se reproducen sexualmente, pero las leyes de la genética mendeliana explican que la variación en la descendencia, nuevamente, es resultado de una mutación dentro de la especie. El concepto de transferencia de genes entre especies no relacionadas no fue considerado como una posibilidad hasta hace relativamente poco tiempo. La transferencia génica horizontal (HGT), también conocida como transferencia lateral de genes, es la transferencia de genes entre especies no relacionadas. Se ha demostrado que el HGT es un fenómeno siempre presente, con muchos evolucionistas postulando un papel importante para este proceso en la evolución, complicando así el modelo de árbol simple. Se ha demostrado que los genes se transmiten entre especies que solo se relacionan de manera lejana usando filogenia estándar, agregando así una capa de complejidad a la comprensión de las relaciones filogenéticas.
Las diversas formas en que el HGT ocurre en los procariotas es importante para comprender las filogenias. Aunque en la actualidad el HGT no se considera importante para la evolución eucariota, el HGT también ocurre en este dominio. Finalmente, como ejemplo de la última transferencia génica, se han propuesto teorías de fusión genómica entre organismos simbióticos o endosimbióticos para explicar un evento de gran importancia: la evolución de la primera célula eucariota, sin la cual los humanos no podrían haber existido.
Transferencia de Genes Horizontal
La transferencia genética horizontal (HGT) es la introducción de material genético de una especie a otra especie por mecanismos distintos a la transmisión vertical de progenitores a crías. Estas transferencias permiten que incluso especies distantemente relacionadas compartan genes, influyendo en sus fenotipos. Se piensa que el HGT es más prevalente en procariotas, pero que solo alrededor del 2% del genoma procariota puede ser transferido por este proceso. Algunos investigadores consideran que tales estimaciones son prematuras: la importancia real de HGT para los procesos evolutivos debe ser vista como un trabajo en progreso. A medida que el fenómeno se investiga más a fondo, puede revelarse que es más común. Muchos científicos creen que el HGT y la mutación parecen ser (especialmente en procariotas) una fuente significativa de variación genética, que es la materia prima para el proceso de selección natural. Estas transferencias pueden ocurrir entre dos especies cualesquiera que compartan una relación íntima (Tabla\(\PageIndex{1}\)).
| Tabla\(\PageIndex{1}\): Resumen de Mecanismos de HGT Procariota y Eucariota | |||
|---|---|---|---|
| Mecanismo | Modo de transmisión | Ejemplo | |
| \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT procariotas y eucariotas” style="height:14px; "> Procariotas | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTMecanismo Procariota y Eucariota” style="height:14px; ">transformation | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTModo de Transmisión Procariota y Eucariota” style="height:14px; ">Captación de ADN | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTEjemplo procariota y eucariota” style="height:14px; ">many procariotas |
| \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariota y Eucariota” style="height:14px; "> | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariota y Eucariota” style="height:14px; ">transducción | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTModo de Transmisión Procariota y Eucariota” style="height:14px; ">bacteriófago (virus) | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTEjemplo procariota y eucariota” style="height:14px; ">bacteria |
| \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariota y Eucariota” style="height:14px; "> | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariota y Eucariota” style="height:14px; ">conjugación | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTModo de Transmisión Procariota y Eucariota” style="height:14px; ">pilus | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTEjemplo procariota y eucariota” style="height:14px; ">many procariotas |
| \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariota y Eucariota” style="height:14px; "> | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariotas y Eucariotas” style="height:14px; ">agentes de transferencia génica | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTModo de Transmisión Procariota y Eucariota” style="height:14px; ">partículas similares a fagos | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariotas y Eucariotas Ejemplo” style="height:14px; ">bacterias moradas sin azufre |
| \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT procariotas y eucariotas” style="height:14px; "> Eucariotas | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTMecanismo Procariota y Eucariota” style="height:14px; ">from food organisms | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTModo de Transmisión Procariota y Eucariota” style="height:14px; ">unknown | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTExample procariota y eucariota” style="height:14px; ">pulgón |
| \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariota y Eucariota” style="height:14px; "> | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTMecanismo Procariota y Eucariota” style="height:14px; ">genes saltantes | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTModo de Transmisión Procariota y Eucariota” style="height:14px; ">transposones | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTEjemplo procariota y eucariota” style="height:14px; ">plantas de arroz y mijo |
| \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariota y Eucariota” style="height:14px; "> | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariotas y Eucariotas” style="height:14px; ">epífitos/parásitos | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTModo de Transmisión Procariota y Eucariota” style="height:14px; ">unknown | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTEjemplo procariota y eucariota” style="height:14px; ">hongos de tejo |
| \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariota y Eucariota” style="height:14px; "> | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGT Procariotas y Eucariotas” style="height:14px; ">de infecciones virales | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTModo de Transmisión Procariota y Eucariota” style="height:14px; "> | \ (\ PageIndex {1}\): Resumen de Mecanismos de HGTEjemplo procariota y eucariota” style="height:14px; "> |
HGT en Procariotas
Se ha demostrado que el mecanismo de HGT es bastante común en los dominios procariotas de Bacterias y Archaea, cambiando significativamente la forma en que se ve su evolución. La mayoría de los modelos evolutivos, como en la Teoría Endosimbionte, proponen que los eucariotas descienden de múltiples procariotas, lo que hace que el HGT sea aún más importante para comprender las relaciones filogenéticas de todas las especies existentes y extintas.
El hecho de que los genes se transfieren entre bacterias comunes es bien conocido por los estudiantes de microbiología. Estas transferencias de genes entre especies son el principal mecanismo por el cual las bacterias adquieren resistencia a los antibióticos. Clásicamente, se ha pensado que este tipo de transferencia ocurre por tres mecanismos diferentes:
- Transformación: el ADN desnudo es captado por una bacteria
- Transducción: los genes se transfieren usando un virus
- Conjugación: el uso de un tubo hueco llamado pilus para transferir genes entre organismos
Más recientemente, se ha descubierto un cuarto mecanismo de transferencia génica entre procariotas. Pequeñas partículas similares a virus llamadas agentes de transferencia génica (GTA) transfieren segmentos genómicos aleatorios de una especie de procariota a otra. Se ha demostrado que los GTA son responsables de los cambios genéticos, a veces con una frecuencia muy alta en comparación con otros procesos evolutivos. El primer GTA se caracterizó en 1974 utilizando bacterias moradas, no azufradas. Estos GTA, que se cree que son bacteriófagos que perdieron la capacidad de reproducirse por sí mismos, transportan piezas aleatorias de ADN de un organismo a otro. La capacidad de los GTA para actuar con alta frecuencia se ha demostrado en estudios controlados con bacterias marinas. Los eventos de transferencia de genes en procariotas marinos, ya sea por GTA o por virus, se han estimado hasta 10 13 por año solo en el Mar Mediterráneo. Se cree que los GTA y los virus son vehículos HGT eficientes con un gran impacto en la evolución procariota.
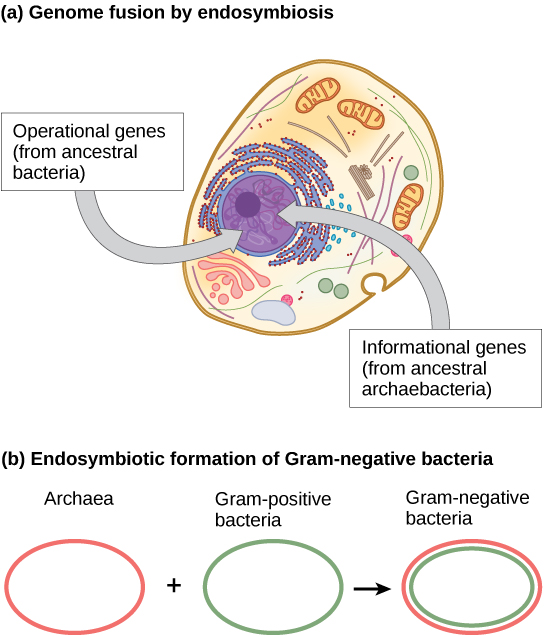
Como consecuencia de este moderno análisis de ADN, la idea de que los eucariotas evolucionaron directamente de Archaea ha caído en desgracia. Si bien los eucariotas comparten muchas características que están ausentes en las bacterias, como la caja TATA (que se encuentra en la región promotora de muchos genes), el descubrimiento de que algunos genes eucariotas eran más homólogos con el ADN bacteriano que el ADN de Archaea hizo que esta idea fuera menos sostenible. Además, la fusión de genomas de Archaea y Bacterias por endosimbiosis se ha propuesto como el evento final en la evolución eucariota.
HGT en Eucariotas
Si bien es fácil ver cómo los procariotas intercambian material genético por HGT, inicialmente se pensó que este proceso estaba ausente en eucariotas. Después de todo, los procariotas no son sino células individuales expuestas directamente a su entorno, mientras que las células sexuales de organismos multicelulares suelen ser secuestradas en partes protegidas del cuerpo. De esta idea se deduce que las transferencias de genes entre eucariotas multicelulares deberían ser más difíciles. En efecto, se piensa que este proceso es más raro en eucariotas y tiene un impacto evolutivo mucho menor que en los procariotas. A pesar de este hecho, el HGT entre organismos distantemente relacionados se ha demostrado en varias especies eucariotas, y es posible que en el futuro se descubran más ejemplos.
En las plantas, se ha observado transferencia génica en especies que no pueden polinizar de manera cruzada por medios normales. Se ha demostrado que los transposones o “genes saltadores” se transfieren entre especies de plantas de arroz y mijo. Además, las especies fúngicas que se alimentan de tejos, de las que deriva el medicamento anticancerígeno TAXOL® de la corteza, han adquirido la capacidad de elaborar taxol ellos mismos, un claro ejemplo de transferencia génica.
En animales, un ejemplo particularmente interesante de HGT ocurre dentro de las especies de pulgones (Figura\(\PageIndex{2}\)). Los pulgones son insectos que varían en color según el contenido de carotenoides. Los carotenoides son pigmentos elaborados por una variedad de plantas, hongos y microbios, y cumplen una variedad de funciones en los animales, quienes obtienen estos químicos de sus alimentos. Los humanos requieren carotenoides para sintetizar la vitamina A, y los obtenemos comiendo frutas y verduras de naranja como zanahorias, albaricoques, mangos y batatas. Por otro lado, los pulgones han adquirido la capacidad de elaborar carotenoides por su cuenta. Según el análisis de ADN, esta capacidad se debe a la transferencia de genes fúngicos al insecto por el HGT, presumiblemente ya que el insecto consumió hongos como alimento. Una enzima carotenoide llamada desaturasa es responsable de la coloración roja observada en ciertos pulgones, y se ha demostrado además que cuando este gen se inactiva por mutación, los pulgones vuelven a su color verde más común (Figura\(\PageIndex{2}\)).

Fusión Genómica y Evolución de Eucariotas
Los científicos creen que lo último en HGT ocurre a través de la fusión genómica entre diferentes especies de procariotas cuando dos organismos simbióticos se vuelven endosimbióticos. Esto ocurre cuando una especie es tomada dentro del citoplasma de otra especie, lo que finalmente da como resultado un genoma compuesto por genes tanto del endosimbionte como del hospedador. Este mecanismo es un aspecto de la Teoría Endosimbionte, la cual es aceptada por la mayoría de los biólogos como el mecanismo por el cual las células eucariotas obtuvieron sus mitocondrias y cloroplastos. Sin embargo, el papel de la endosimbiosis en el desarrollo del núcleo es más controvertido. Se cree que el ADN nuclear y mitocondrial son de origen evolutivo diferente (separado), con el ADN mitocondrial derivado de los genomas circulares de bacterias que fueron engulladas por antiguas células procariotas. El ADN mitocondrial puede considerarse como el cromosoma más pequeño. Curiosamente, el ADN mitocondrial se hereda solo de la madre. El ADN mitocondrial se degrada en los espermatozoides cuando el esperma se degrada en el óvulo fertilizado o en otros casos cuando las mitocondrias localizadas en el flagelo del espermatozoide no logran ingresar al óvulo.
En la última década, el proceso de fusión genómica por endosimbiosis ha sido propuesto por James Lake del Instituto de Astrobiología UCLA/NASA como responsable de la evolución de las primeras células eucariotas (Figura\(\PageIndex{3}\) a). Mediante el análisis de ADN y un nuevo algoritmo matemático llamado reconstrucción condicionada (RC), su laboratorio propuso que las células eucariotas se desarrollaran a partir de una fusión génica endosimbiótica entre dos especies, una Archaea y la otra una Bacteria. Como se mencionó, algunos genes eucariotas se asemejan a los de Archaea, mientras que otros se parecen a los de Bacterias Un evento de fusión endosimbiótica, como el que Lake ha propuesto, explicaría claramente esta observación. Por otro lado, este trabajo es nuevo y el algoritmo CR es relativamente infundado, lo que hace que muchos científicos se resistan a esta hipótesis.
Un trabajo más reciente de Lake (Figura\(\PageIndex{3}\) b) propone que las bacterias gramnegativas, que son únicas dentro de su dominio en el sentido de que contienen dos membranas bicapa lipídicas, en efecto, resultaron de una fusión endosimbiótica de especies arqueales y bacterianas. La doble membrana sería un resultado directo de la endosimbiosis, con el endosimbionte recogiendo la segunda membrana del huésped a medida que se internalizaba. Este mecanismo también se ha utilizado para explicar las membranas dobles que se encuentran en mitocondrias y cloroplastos. El trabajo de Lake no está exento de escepticismo, y las ideas aún se debaten dentro de la comunidad de ciencias biológicas. Además de la hipótesis de Lake, hay varias otras teorías en competencia sobre el origen de los eucariotas. ¿Cómo evolucionó el núcleo eucariota? Una teoría es que las células procariotas produjeron una membrana adicional que rodeaba el cromosoma bacteriano. Algunas bacterias tienen el ADN encerrado por dos membranas; sin embargo, no hay evidencia de un nucleolo o poros nucleares. Otras proteobacterias también tienen cromosomas unidos a la membrana. Si el núcleo eucariota evolucionara de esta manera, esperaríamos que uno de los dos tipos de procariotas estuviera más estrechamente relacionado con los eucariotas.
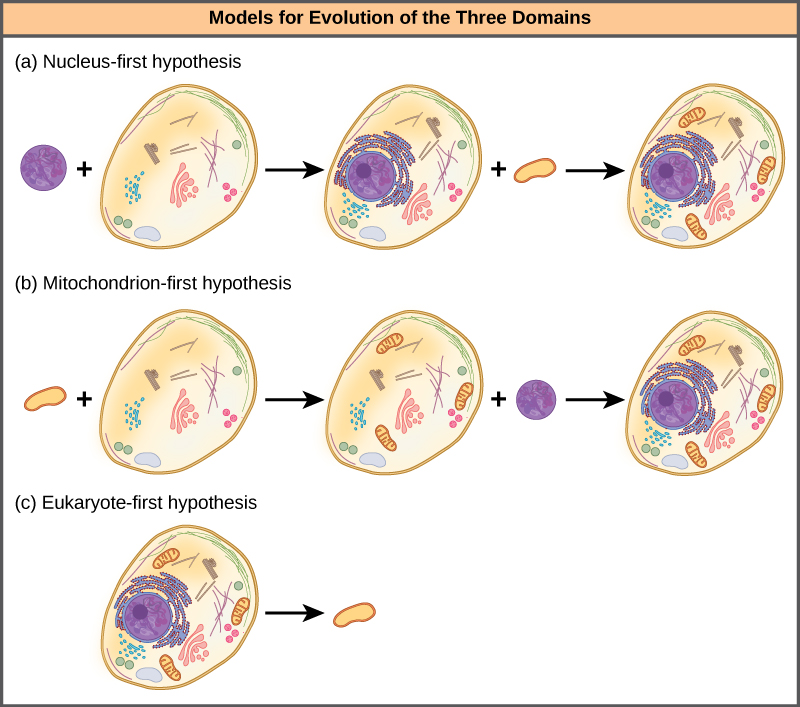
La hipótesis del núcleo primero propone que el núcleo evolucionó primero en procariotas (Figura\(\PageIndex{4}\) a), seguido de una fusión posterior del nuevo eucariota con bacterias que se convirtieron en mitocondrias. La hipótesis mitocondria-primera propone que las mitocondrias se establecieron por primera vez en un huésped procariota (Figura\(\PageIndex{4}\) b), que posteriormente adquirió un núcleo, por fusión u otros mecanismos, para convertirse en la primera célula eucariota. La hipótesis eucariota primero propone que los procariotas realmente evolucionaron a partir de eucariotas al perder genes y complejidad (Figura\(\PageIndex{4}\) c). Todas estas hipótesis son comprobables. Solo el tiempo y más experimentación determinarán qué hipótesis es mejor apoyada por los datos.
Modelos web y de red
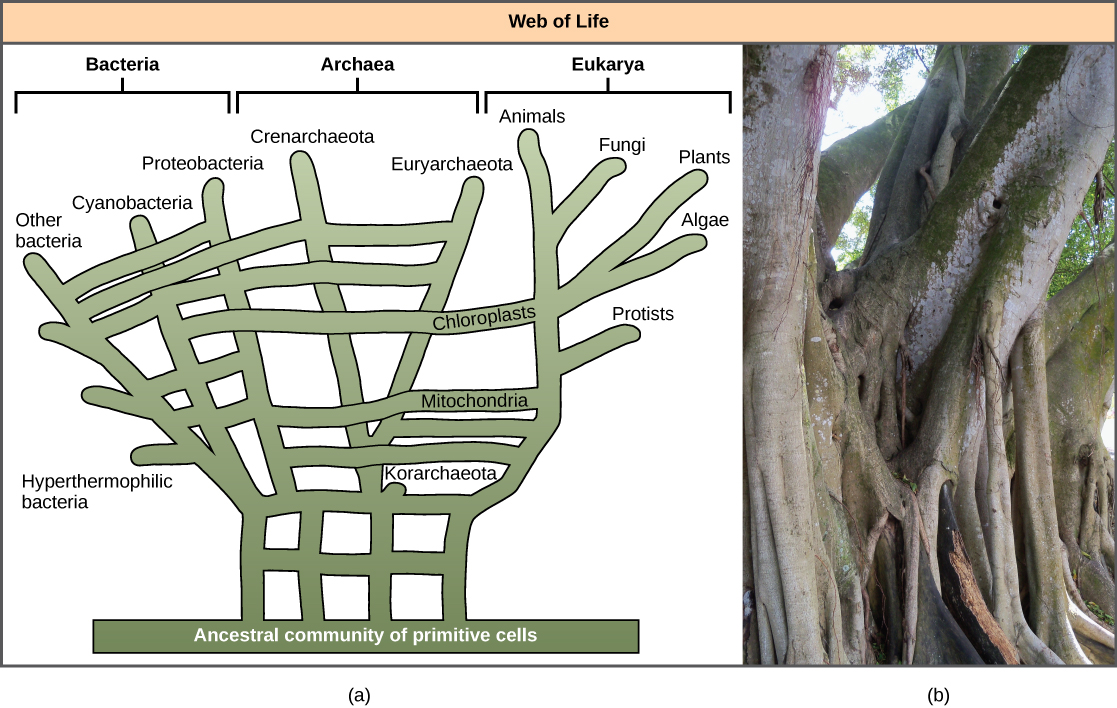
El reconocimiento de la importancia del HGT, especialmente en la evolución de los procariotas, ha provocado que algunos propongan abandonar el modelo clásico de “árbol de la vida”. En 1999, W. Ford Doolittle propuso un modelo filogenético que se asemeja más a una red o red que a un árbol. La hipótesis es que los eucariotas evolucionaron no de un solo ancestro procariota, sino de un charco de muchas especies que compartían genes por mecanismos HGT. Como se muestra en la Figura\(\PageIndex{5}\) a, algunos procariotas individuales fueron los encargados de transferir las bacterias que causaron el desarrollo mitocondrial a los nuevos eucariotas, mientras que otras especies transfirieron las bacterias que dieron origen a los cloroplastos. A este modelo se le suele llamar la “red de la vida”. ” En un esfuerzo por salvar la analogía del árbol, algunos han propuesto usar el árbol Ficus (Figura\(\PageIndex{5}\) b) con sus múltiples troncos como filogenético para representar un papel evolutivo disminuido para HGT.
Modelos de anillo de vida
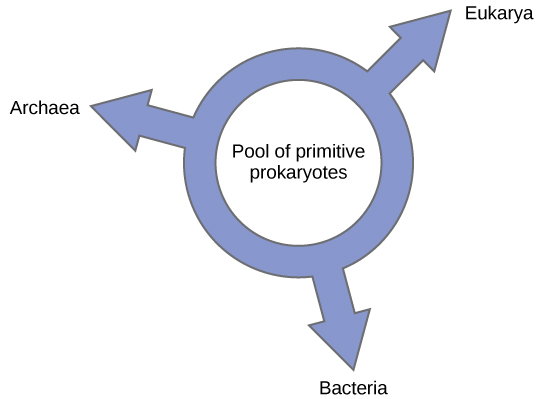
Otros han propuesto abandonar cualquier modelo de filogenia similar a un árbol en favor de una estructura de anillo, el llamado “anillo de vida” (Figura\(\PageIndex{6}\)); un modelo filogenético donde los tres dominios de la vida evolucionaron a partir de un charco de procariotas primitivos. Lake, nuevamente usando el algoritmo de reconstrucción condicionada, propone un modelo similar a un anillo en el que especies de los tres dominios, Archaea, Bacteria y Eucarya, evolucionaron a partir de un solo grupo de procariotas de intercambio de genes. Su laboratorio propone que esta estructura es la más adecuada para los datos de extensos análisis de ADN realizados en su laboratorio y que el modelo de anillo es el único que toma adecuadamente en cuenta el HGT y la fusión genómica. Sin embargo, otros filogenetistas siguen siendo altamente escépticos de este modelo.
En resumen, el modelo de “árbol de la vida” propuesto por Darwin debe ser modificado para incluir HGT. ¿Significa esto abandonar completamente el modelo arbóreo? Incluso Lake sostiene que se deben hacer todos los intentos para descubrir alguna modificación del modelo de árbol para permitir que se ajuste con precisión a sus datos, y solo la incapacidad de hacerlo influirá a la gente hacia su propuesta de anillo.
Esto no significa que un árbol, una red o un anillo se correlacionen completamente con una descripción precisa de las relaciones filogenéticas de la vida. Una consecuencia del nuevo pensamiento sobre los modelos filogenéticos es la idea de que la concepción original de Darwin del árbol filogenético es demasiado simple, pero tenía sentido a partir de lo que se conocía en ese momento. Sin embargo, la búsqueda de un modelo más útil avanza: cada modelo sirve como hipótesis a probar con la posibilidad de desarrollar nuevos modelos. Así avanza la ciencia. Estos modelos se utilizan como visualizaciones para ayudar a construir relaciones evolutivas hipotéticas y comprender la enorme cantidad de datos que se analizan.
Resumen
El árbol filogenético, utilizado por primera vez por Darwin, es el modelo clásico de “árbol de la vida” que describe las relaciones filogenéticas entre especies, y el modelo más común utilizado hoy en día. Las nuevas ideas sobre HGT y la fusión genómica han hecho que algunos sugieran revisar el modelo para asemejarse a redes o anillos.
Referencias
OpenStax, Biología. OpenStax CNX. 25 de junio de 2020. https://cnx.org/contents/GFy_h8cu@10.137:noBcfThl@7/Understanding-Evolution.