
5.6: Traducción
- Page ID
- 53518

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
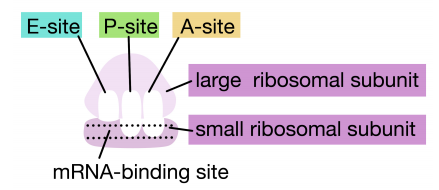
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)La traducción es el proceso por el cual se utiliza la información en los ARNm para dirigir la síntesis de proteínas. Como has aprendido en biología introductoria, en células eucariotas, este proceso se lleva a cabo en el citoplasma de la célula, por grandes máquinas ARN-proteína llamadas ribosomas. Los ribosomas contienen ARN ribosómicos (ARNr) y proteínas. Las proteínas y ARNr se organizan en dos subunidades, una grande y una pequeña. La subunidad grande tiene una actividad enzimática, conocida como peptidil-transferasa, que hace que los enlaces peptídicos que unen los aminoácidos para formar un polipéptido. Las subunidades pequeñas y grandes se ensamblan en el ARNm en su extremo 5' para iniciar la traducción. Los ribosomas funcionan uniéndose a los ARNm y sujetándolos de una manera que permita que los aminoácidos codificados por el ARN se unan secuencialmente para formar un polipéptido.
La secuencia de un ARNm especifica directamente la secuencia de aminoácidos en la proteína que codifica. Cada aminoácido en la proteína está especificado por una secuencia de 3 bases llamada codón en el ARNm. Por ejemplo, el aminoácido triptófano está codificado por la secuencia 5'UGG3' en un ARNm. Dado que hay 4 bases en el ARN, el número de diferentes combinaciones de 3 bases que son posibles es\(4^3\), o 64. Sin embargo, solo hay 20 aminoácidos que se utilizan en la construcción de proteínas. Esta discrepancia en el número de codones posibles y el número real de aminoácidos que especifican se explica por el hecho de que el mismo aminoácido puede ser especificado por más de un codón. De hecho, con la excepción de los aminoácidos metionina y triptófano, todos los demás aminoácidos están codificados por múltiples codones. La figura anterior muestra los codones que se utilizan para cada uno de los veinte aminoácidos.
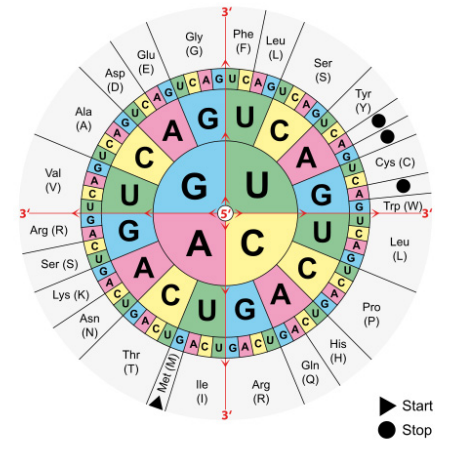
Tres de los 64 codones se conocen como codones de terminación o terminación y como su nombre indica, indican el final de una secuencia codificante de proteína. El codón para metionina, AUG, se usa como codón de inicio, o iniciación.
Este ingenioso sistema se utiliza para dirigir el ensamblaje de una proteína de la misma manera que podrías ensartar cuentas de colores en un orden particular usando instrucciones que usaban símbolos como 111 para una cuenta roja, seguido de 222 para una cuenta verde, 333 para amarillo, y así sucesivamente, hasta llegar a 000, indicando que debe dejar de encordar cuentas.
Si bien los ribosomas son literalmente las fábricas de proteínas que unen aminoácidos juntos usando las instrucciones en los ARNm, otra clase de moléculas de ARN, los ARN de transferencia (ARNt) también son necesarios para la traducción. Los ARN de transferencia (ver figura, izquierda) son moléculas de ARN pequeñas, de aproximadamente 75-80 nucleótidos de longitud, que funcionan para 'interpretar' las instrucciones en el ARNm durante la síntesis de proteínas. En términos de la analogía de cuentas anterior, alguien, o algo, tiene que poder traer una cuenta roja cuando las instrucciones indican 111, y una cuenta verde cuando las instrucciones dicen 222. A diferencia de un humano, que puede elegir una cuenta roja cuando 111 está presente en las instrucciones, ni los ribosomas ni los ARNt pueden pensar. El sistema, por lo tanto, se basa, como tantos procesos en las células, únicamente en el reconocimiento molecular.
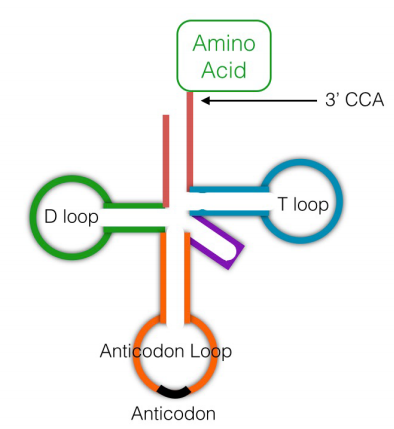
Un ARN de transferencia dado es específico para un aminoácido particular. Está unido covalentemente a este aminoácido en su extremo 3' por una enzima llamada aminoacil ARNt sintetasa. Existe una aminoacil ARNt sintetasa específica para cada aminoácido. Se dice que un ARNt con un aminoácido unido a él está cargado. Otra región del ARNt tiene una secuencia de 3 bases, el anticodón, que es complementario al codón para el aminoácido que lleva. Cuando el ARNt encuentra el codón para su aminoácido en el ARN mensajero, el anticodón se emparejará por bases con el codón, y el aminoácido unido a él se llevará al ribosoma para agregarlo a la cadena proteica en crecimiento.
Con una idea de los diversos componentes necesarios para la traducción ahora podemos echar un vistazo al proceso de síntesis de proteínas. Los principales pasos en el proceso son similares en procariotas y eucariotas. Como ya señalamos, los ribosomas se unen a los ARNm y facilitan la interacción entre los codones en el ARNm y los anticodones en los ARNt cargados.
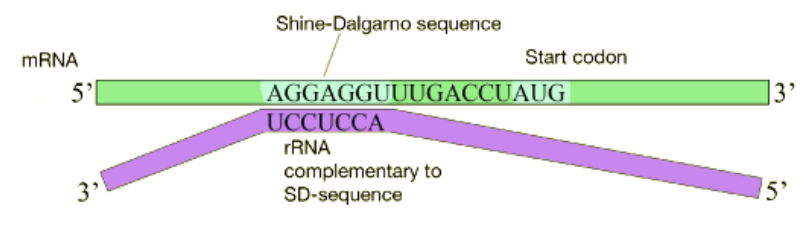
En las células bacterianas, la traducción se acopla con la transcripción y comienza incluso antes de que el ARNm se haya sintetizado completamente. ¿Cómo reconoce el ribosoma y se une al ARNm? Muchos ARNm bacterianos portan una secuencia corta rica en purinas conocida como el sitio Shine-Dalgarno aguas arriba del codón de inicio AUG, como se muestra en la siguiente figura. Esta secuencia es reconocida y unida por una secuencia complementaria en el ARNr 16S que forma parte de la subunidad ribosómica pequeña como se muestra anteriormente. Debido a que el sitio Shine-Dalgarno sirve para reclutar y unir el ribosoma, también se le conoce como el sitio de unión al ribosoma o RBS.

Una variación de este proceso de ensamblaje de ribosomas opera en células eucariotas. Ya sabemos que en las células eucariotas, los ARNm procesados son enviados desde el núcleo al citoplasma.
Las subunidades pequeñas y grandes de los ribosomas, cada una compuesta por ARNs y proteínas característicos, se encuentran en el citoplasma y se ensamblan sobre los ARNm para formar ribosomas completos que llevan a cabo la traducción.
La síntesis de proteínas en eucariotas comienza con la unión de la subunidad pequeña del ribosoma al extremo 5' del ARNm. El ensamblaje de la maquinaria de traducción comienza con la unión de la subunidad ribosómica pequeña a la tapa de 7-metil guanosina en el extremo 5' de un ARNm. Mientras tanto, el ARNt iniciador se empareja con el codón de inicio. (Recordemos que el codón de inicio es AUG, y codifica metionina. El ARNt iniciador porta el aminoácido metionina). La subunidad grande del ribosoma se une luego al complejo, que ahora está listo para iniciar la síntesis de proteínas.
Los ribosomas tienen dos sitios para unirse a ARNt cargados, cada uno de los cuales se posiciona para hacer que dos codones adyacentes en el ARNm estén disponibles para la unión por ARNt. El codón de iniciación ocupa el primero de estos sitios ribosómicos, el sitio P. El anticodón complementario a éste está en el ARNt iniciador, que aporta el primer aminoácido de la proteína. Esta fase inicial de traducción se llama iniciación y requiere la ayuda de factores proteicos llamados factores de iniciación.
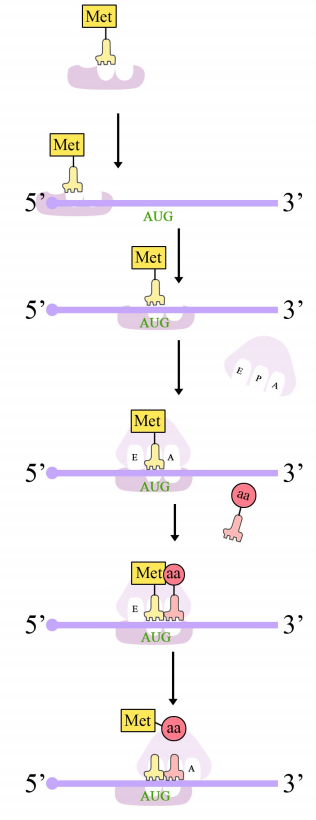
El segundo codón del ARNm se posiciona adyacente al segundo sitio en el ribosoma, el sitio A. Aquí es donde se une el ARNt que porta el aminoácido especificado por el segundo codón. La unión de aminoacil ARNt al sitio A está mediada por proteínas llamadas factores de elongación y requiere el aporte de energía. Una vez que los ARNt cargados apropiados se han “acoplado” en los codones por apareamiento de bases entre el anticodón en el ARNt y el codón en el ARNm, el ribosoma une los aminoácidos portados por los dos ARNt haciendo un enlace peptídico (ver figura a la derecha).
Curiosamente, la formación del enlace peptídico es catalizada por un ARN catalítico (el ARNr 23S en procariotas) en lugar de por una enzima proteica.
Esta y las etapas posteriores en la síntesis del polipéptido se denominan fase de elongación de la traducción. Una vez que los dos primeros aminoácidos están unidos, el primer ARNt se disocia y se mueve fuera del sitio P y al sitio E, o Exit. El segundo ARNt luego se mueve al sitio P, dejando el sitio A para el ARNt correspondiente al siguiente codón.
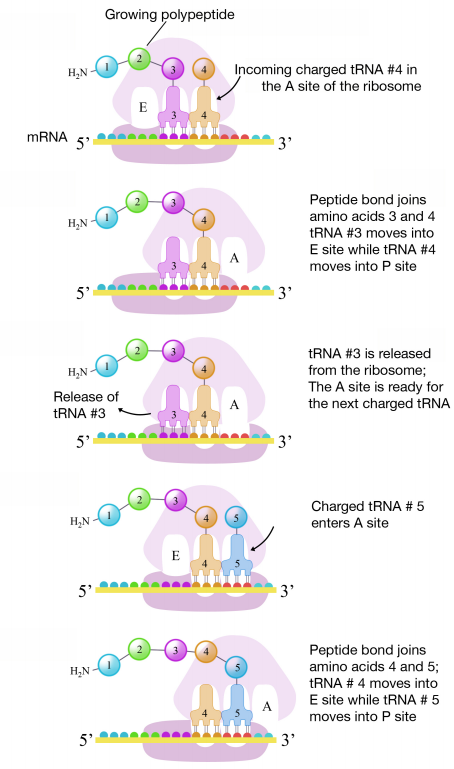
El proceso se repite hasta que el codón de terminación está en el sitio A. En este punto, un factor de liberación se une al sitio A, agrega una molécula de agua al polipéptido en el sitio P, y libera el polipéptido completo del ribosoma, que a su vez, luego se disocia en subunidades.
Como se describe en el Capítulo 3, los polipéptidos elaborados de esta manera se pliegan luego en sus formas tridimensionales, se modifican postraduccionalmente y se entregan a los compartimentos celulares apropiados para llevar a cabo sus funciones.