
4.4: Ciclo de Vida del Eucariote Multicelular
- Page ID
- 55327

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Origen de la Muerte
Las células no siempre se separan después de la mitosis, sino que a veces permanecen juntas para formar organismos multicelulares. Esto aumenta su tamaño, y por lo tanto proporciona una defensa contra los depredadores. Desafortunadamente, no es posible simplemente aumentar el tamaño de la célula porque la célula realmente grande tendrá menos superficie (en relación con el volumen), por lo tanto tendrá múltiples dificultades con la fotosíntesis, respiración y otros procesos que se relacionan con la superficie de la célula. Pero muchas células juntas harán que la superficie sea lo suficientemente grande (Figura\(\PageIndex{1}\)). El organismo multicelular tiene dos modos de crecimiento: escalar el cuerpo y multiplicar las células.

Figura\(\PageIndex{1}\) Origen de la multicelularidad. No es factible solo agrandar la celda, la superficie es demasiado pequeña. Pero si las células no se separan después de la mitosis, podrían formar el cuerpo que es lo suficientemente grande como para escapar de los depredadores. Esto también proporciona un nuevo modo de crecimiento y posibilidad de la división del trabajo (células coloreadas).
La multicelularidad permite que estas células también dividan el trabajo de parto y cooperen. Esto es sumamente importante para la evolución futura.
Las células en el cuerpo multicelular no están conectadas para siempre. En ocasiones, una o pocas células escapan y comienzan un nuevo cuerpo. Este cuerpo será copia exacta (clon) del anterior (reproducción vegetativa). También es posible que cuando estas “células escapadas” vayan por la ruta diferente: se conviertan en “delegados sexuales”, gametos. Todos los gametos quieren singamia, y estas células buscarán al compañero de la misma especie pero con otro genotipo. En caso de heterogamia y oogamia, es fácil de reconocer porque los géneros proporcionarán una pista: el macho buscará a la hembra. En caso de isogamia, los gametos buscan la pareja con diferentes proteínas de superficie. Después de que finalmente se aparean, aparece una célula diploide (cigoto). El cigoto puede invernar y luego dividirse meióticamente. Este es el ciclo de vida más simple del organismo multicelular (Figura\(\PageIndex{2}\)), bastante similar al ciclo discutido anteriormente para el organismo unicelular.

Figura Ciclo de vida\(\PageIndex{2}\) más antiguo del organismo multicelular. El cigoto no crece, se divide meióticamente. Las células somáticas (“grises”) van a morir, solo las células germinales transfieren su ADN a las generaciones futuras.
Sin embargo, con frecuencia el cigoto comienza a crecer y dividirse mitóticamente, haciendo que el cuerpo diploide. Hay dos razones para hacer cuerpo multicelular a partir del cigoto sin meiosis: (a) porque en lata y (b) porque diploide es mejor. “Puede” porque el cigoto ya contiene programa de ADN sobre cómo construir el cuerpo multicelular. Por qué es mejor diploide, explicado en la siguiente sección.
Si el organismo multicelular consiste en células diploides (\(2n\)), usaremos el término neutro diplont. Los organismos multicelulares con células haploides (\(n\)) son haplontes.
“Células escapadas”, “delegados sexuales”, o células madre de gametos de lo anterior es una primera etapa de la división del trabajo cuando las células se están separando en dos tipos, células germinales y células somáticas. Las células somáticas son aquellas que eventualmente morirán, pero las células germinales son capaces de dar descendencia. Tener células germinales no es absolutamente necesario para los organismos multicelulares, pero la mayoría de ellos tienen líneas germinales bien separadas. Así, el origen de la muerte está directamente relacionado con esta separación: las células somáticas no son necesarias para las generaciones futuras. Los organismos unicelulares son potencialmente inmortales, y lo mismo son las células cancerosas que también escapan del organismo (pero no pueden hacer la nueva).
El ciclo de vida del organismo multicelular podría describirse a partir del haplonte (Figura\(\PageIndex{3}\)). Cuando las condiciones ambientales son favorables, tiene reproducción vegetativa. Una variante de la reproducción vegetativa es que la célula (mitosporas) se separa de un haplonte, luego se divide en más células y se convierte en un nuevo haplonte. En ocasiones, los trozos enteros se separan y se convierten en nuevos haplontes. Cuando cambian las condiciones, el haplonte puede iniciar la reproducción sexual: la singamia. En la singamia, un gameto se separa del haplonte y se une con un gameto de otro haplonte. Juntos, los gametos forman un cigoto. Este cigoto podría ir directo a la meiosis (como ocurre en los eucariotas unicelulares) pero con mayor frecuencia, el cigoto crecerá, se dividirá mitóticamente y finalmente se convertirá en diplonte. Este diplonte puede ser superficialmente casi idéntico al haplonte pero cada célula del mismo contiene núcleo diploide (cada cromosoma tiene un par). Diplont (similar al haplonte) puede reproducirse vegetativamente (hacer clones): la célula se separa de un diplonte, luego se divide mitóticamente en más células y se convierte en un nuevo diplonte.
El diplonte también es capaz de reproducción asexual: podría haber una célula que se separa de un diplonte y se divide con meiosis creando cuatro esporas, cada una de ellas crecerá en haplonte.
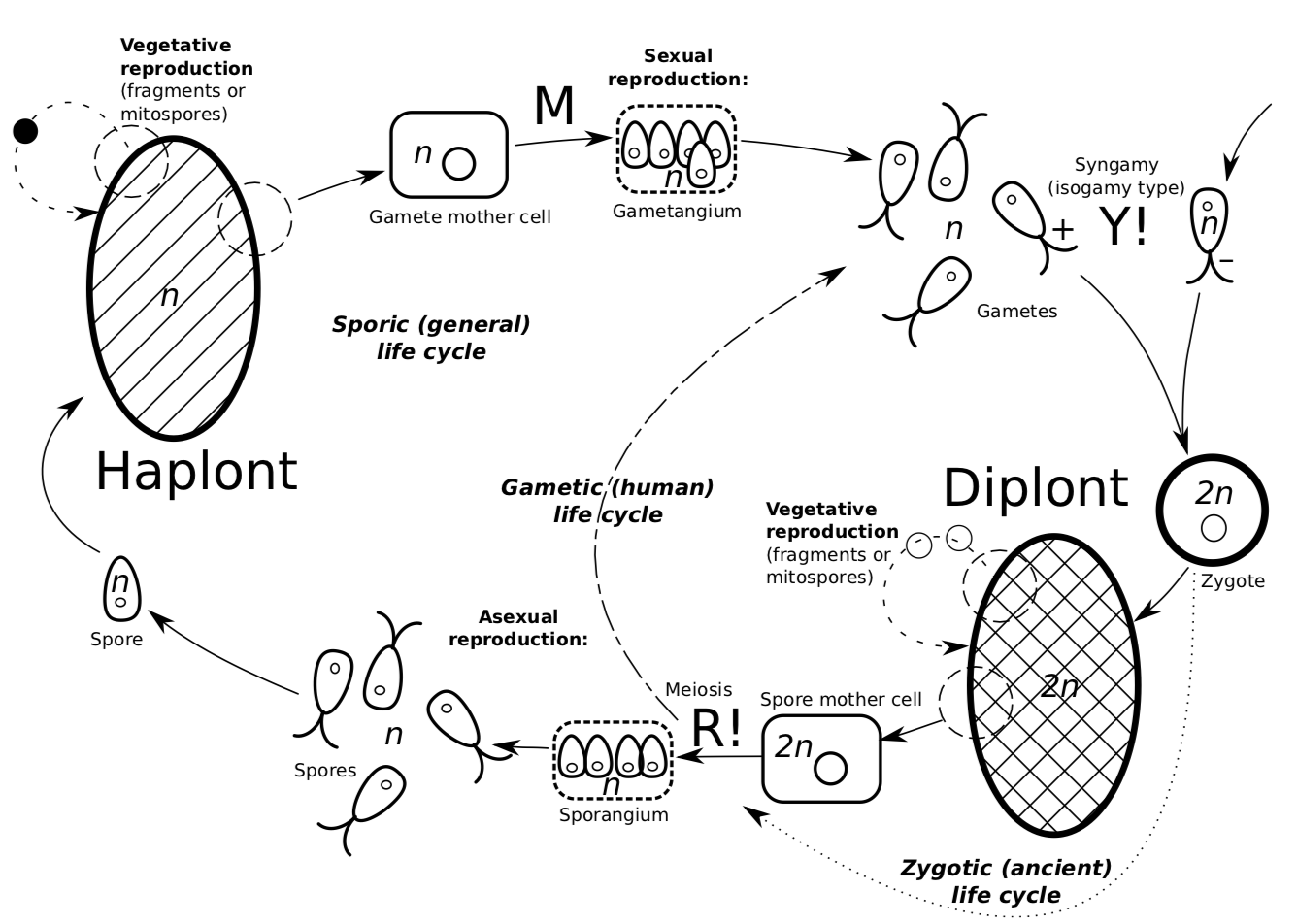
Figura Ciclo de vida\(\PageIndex{3}\) general. La parte haploide está a la izquierda, diploide a la derecha, la singamia en la parte superior, la meiosis en la parte inferior. La letra “M” se usa para etiquetar la mitosis.
Ciclos de vida esporicos, cigóticos y góticos
El ciclo de vida descrito anteriormente es el ciclo de vida esporico (Figura\(\PageIndex{4}\)). Los organismos con ciclo de vida esporico tienen tanto diplont como haplontes, igual o desigualmente desarrollados.

Figura Ciclo de vida\(\PageIndex{4}\) esporico. Visión general. La parte haploide está a la izquierda, diploide a la derecha, la singamia en la parte superior, la meiosis en la parte inferior.
En total, existen tres tipos de ciclos de vida: esporico, cigótico, que es el más parecido al unicelular y el más primitivo; y el gético, que es utilizado por animales y algunos protistas (Figura\(\PageIndex{5}\)). El ciclo de vida cigótico comienza con la singamia y pasa a la meiosis. No tiene diplont. El ciclo de vida gametico va de la meiosis a la singamia. No tiene haplonte.
Los protistas tienen los tres tipos de ciclos de vida mientras que los grupos superiores tienen solo uno. Los animales exhiben ciclo gamético, mientras que las plantas\(_2\) retuvieron el ciclo esporico más primitivo.
Evolución de los Ciclos de Vida
La diferencia más llamativa entre los ciclos de vida unicelulares y multicelulares es que el cigoto del organismo multicelular puede comenzar a formar cuerpo diploide (diplonte) que a veces es visualmente casi idéntico al haplonte. Esto se debe a que en la perspectiva evolutiva, los diplontes son “mejores” que los haplontes. La situación frecuente de dominancia génica permite que sólo una variante (alelo) del gen funcione, que puede salvar al organismo de mutaciones letales. Un mayor número de genes podría ayudar a producir más proteínas. Una tercera razón es que los genomas de los diplomados son más diversos. Un gen puede ser capaz de soportar un grupo de condiciones, y la otra variante puede tener un conjunto diferente de condiciones posibles. Por lo tanto, diplont es capaz de aprovechar las capacidades de ambas variantes genéticas.
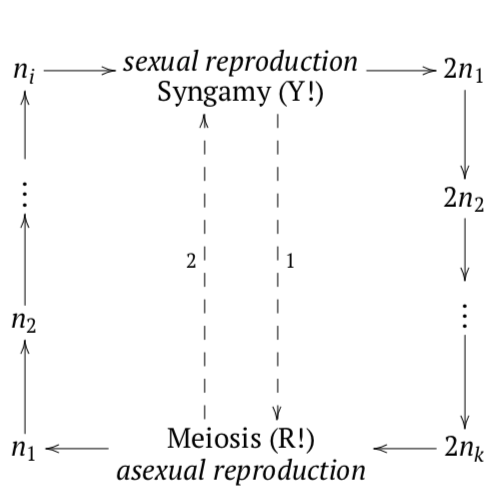
Figura\(\PageIndex{5}\) La evolución de los ciclos de vida (las flechas verdes representan cinco transiciones evolutivas) de cigótico unicelular a gamético multicelular a través de diferentes variantes de ciclos esporicos.
Como consecuencia, la evolución de los ciclos de vida pasa del cigótico (similar al unicelular) al ciclo esporico (Figura\(\PageIndex{6}\)), y luego a la dominación cada vez más expresada del diplonte, y finalmente a la reducción completa del ciclo de vida haplonte, gético. Todavía es una pregunta abierta cómo evolucionaron los protistas cigóticos hacia el lado esporico. Lo más probable es que el cigoto (que por definición es diploide) no quiso dividirse meióticamente. En cambio, crece (lo que se ve en algunos protistas) y se divide mitóticamente, dando a luz al diplont. Así comenzó el primer ciclo esporico. El último paso de esta cadena evolutiva fue una reducción completa del haplonte: después de la meiosis, las esporas fueron reemplazadas por gametos que inmediatamente pasan a la singamia.
Ciclo de Vida de Vegetabilia
Los ancestros de Vegetabilia (plantas\(_2\)) fueron algas verdes con ciclo de vida cigótico. Se podría imaginar que su cigoto comenzó a crecer porque estos organismos habitaban aguas poco profundas y quieren que sus esporas se distribuyan con un viento. Una forma de que esto suceda es tener las esporas en el tallo de la planta. Esta es probablemente la razón

Figura\(\PageIndex{6}\) La evolución de los ciclos de vida (las flechas verdes representan cinco transi- ciones evolutivas) de cigótico unicelular a gamético multicelular a través de diferentes variantes de ciclos esporicos.
de crecimiento de cigotos: los diplontes primordiales de las plantas\(_2\) fueron simplemente esporangios, estructuras portadoras de esporas. Entonces comenzaron a aparecer los beneficios de la condición diploide descrita anteriormente, y estas plantas primitivas se adentraron en el camino de reducción de haplones. Sin embargo, algunos Vegetabilia (hepáticas, musgos y hornworts), todavía tienen dominación haplontes. Esto probablemente se deba a que sus haplones son poiquilohídricos (se explica en los próximos capítulos), adaptación que es beneficiosa para plantas pequeñas.
El ciclo de vida de las plantas\(_2\) es esporico, pero la tradición científica utiliza nombres relacionados con plantas para las etapas. El ciclo (Figura\(\PageIndex{7}\)) comienza con un diplonte llamado esporofito, el cual produce esporas. El esporofito lleva un esporangio, dentro del cual la célula madre de esporas utiliza la meiosis para hacer esporas. Las esporas germinan y se convierten en haplontes llamados gametofitos. El gametofito produce gametos, específicamente un espermatozoide (o simplemente “espermatozoides”) y un ovocito (óvulo). Estos gametos se desarrollan en órganos especiales: gametangios. El gametangio que contiene gametos masculinos (espermatozoides) se llama anteridio, y el gametangio femenino es archegonio, el último normalmente contiene solo un óvulo (ovocito).
Por singamia (oogamia en este caso), los dos gametos forman un cigoto. A continuación, un esporófito joven crece en el gametofito, y finalmente, el ciclo vuelve a comenzar. Nuevamente, el esporofito de Vegetabilia inicia su vida como parásito en el gametofito. Incluso las plantas con flores tienen esta etapa llamada embrión. Tal vez, es por ello que el gametofito de las plantas nunca se\(_2\) ha reducido por completo para transformar su ciclo en gametic. Incluso en los linajes de plantas más avanzados, sus gametofitos masculinos (que solo producen espermatozoides) y femeninos tienen mínimo 3 y 4 células, respectivamente, ¡pero no 0!

Figura Ciclo de\(\PageIndex{7}\) vida de las plantas terrestres. El color rojo se utiliza para las innovaciones, comparando con el esquema de ciclo de vida anterior (general).