
1.24: Los múltiples roles del movimiento y la difusión de materiales en la biología vegetal
- Page ID
- 56422

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Además de necesitar materiales, los organismos necesitan mover materiales: los materiales deben trasladarse desde donde se adquieren o sintetizan a otros lugares donde se utilizan; los materiales pueden necesitar ser transportados a sitios de almacenamiento y también recuperados de los sitios de almacenamiento; las señales químicas pueden necesitar moverse de un lugar de sensación a un lugar de respuesta.
Los materiales se mueven de tres formas básicas en los organismos, dos de las cuales ocurren también en sistemas no vivos: (1) los materiales se mueven por difusión, lo que es consecuencia del hecho de que todas las moléculas a una temperatura superior al cero absoluto (¡es decir, todas las moléculas!) se mueven de manera térmica aleatoria; (2) los materiales fluidos (es decir, gases y líquidos) se mueven por flujo másico, de alta presión a baja siempre que exista una vía abierta que permita que el fluido se mueva; (3) los materiales se mueven como resultado de interacciones químicas únicas de entidades biológicas, las llamadas proteínas motoras, que puede usar energía química para hacer trabajo físico (es decir, empujar o tirar de una molécula, aplicar fuerza a una distancia) o los motores rotativos de flagelos que crean un movimiento de rotación que “corre sobre” el movimiento de protones por un gradiente electroquímico. Las proteínas motoras son altamente significativas dentro de las células donde pueden producir flujo citoplásmico, un proceso que es significativo para las células más grandes porque la difusión es ineficaz excepto en distancias muy cortas. Las células más grandes que se encuentran en algunas de las algas, y especialmente en las células coenocíticas/sifonáceas, dependen en gran medida de la capacidad de las proteínas motoras para transportar material dentro de la célula. Pero para este curso, nos interesa más la capacidad de los organismos multicelulares más grandes, especialmente las plantas, para mover materiales dentro del organismo, proceso que se logra mediante una combinación de difusión y flujo de presión.
TEMAS
- Difusión
- Un modelo simple pero insuficiente
- OSMOSIS: la difusión de solventes, incluida el agua
- Efectos de la presión sobre la difusión
- Combinando los efectos de la pureza y la presión
- Plantas y Hongos utilizan manipulaciones de presión y ósmosis de múltiples vías
- Estructuralmente
- Crecimiento
- Movimientos de celdas de guardia
- Movimientos de folleto
- Transporte de larga distancia
- En el floema
- En el xilema
- los patrones
- los mecanismos
- el problema de la cavitación y las compensaciones en la anatomía del xilema
- casos raros de xilema presurizado
Difusión
Un modelo simple pero insuficiente
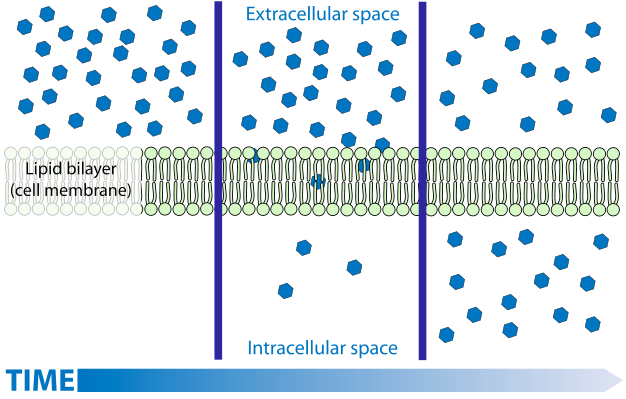
La difusión es un proceso familiar para la mayoría. Si bien la difusión de gases y solutos se entiende fácilmente por modelos simplistas, la comprensión de la difusión de líquidos, en particular el agua, es mucho más desafiante y a menudo se ve confusa por la aplicación imprecisa de la terminología. La difusión de gases y solutos se describe como un movimiento espontáneo de regiones de mayor concentración a regiones de menor concentración. La explicación de este proceso espontáneo se vincula fácilmente a la teoría cinética: las moléculas se mueven de manera aleatoria debido a la energía térmica. Como consecuencia de este movimiento aleatorio, hay un flujo neto de materia desde lugares donde hay más moléculas (es decir, concentraciones más altas) a regiones donde hay menos moléculas (concentraciones más bajas). Si se trata de mezclas (un gas con más de un componente), cada componente se moverá independientemente de los demás.
La difusión de agua líquida (ósmosis) NO siempre es de 'alta concentración a baja'
Sin embargo, este modelo (explicación) no se aplica fácilmente al agua líquida con solutos, o a soluciones en general, en gran parte porque la idea de 'concentración' varía La concentración no es un término tan preciso como uno podría pensar, puede expresarse de varias maneras diferentes (concentración en masa, concentración numérica, molaridad, molalidad, fracción molar). En el modelo simplista de difusión, la medida más apropiada parecería ser (número) concentración (número de moléculas por unidad de volumen) ya que el movimiento aleatorio movería las moléculas desde donde tienen más moléculas por unidad de volumen hasta donde tienen menos moléculas por unidad de volumen. Sin embargo, al considerar el disolvente (no el soluto), la concentración numérica no es un predictor preciso de difusión. Para el agua y para la mayoría de los disolventes, la concentración numérica del disolvente cambia muy poco a medida que se agregan solutos, sin embargo, la adición de solutos puede tener una influencia sustancial en la difusión del disolvente. Y si bien el volumen de agua cambia poco a medida que se agregan solutos, la medida en que sí cambia varía con diferentes solutos, sin embargo, el efecto sobre la difusión del agua no está controlado por el soluto específico agregado sino solo en la cantidad de soluto (número de partículas) que se agregó.Considere que si bien la adición de la mayoría solutos hace que el agua aumente ligeramente en volumen (la concentración numérica baja —el mismo número de moléculas de agua están ahora en un volumen mayor), la adición de algunos solutos puede hacer que el agua se contraiga (es decir, la concentración numérica sube— el mismo número de moléculas de agua ahora están en un volumen menor ). Si la (número) concentración de agua es lo que dirige su movimiento uno esperaría que diferentes solutos tuvieran diferentes efectos en la difusión del líquido, dependiendo de cuánto provocaron que la solución cambiara de volumen (y concentración numérica). Y en realidad se esperaría que algunas soluciones (las que hacen que el agua se encoja) tengan agua difundida de la solución en agua pura. ¡Esto nunca sucede! Manteniendo constantes todos los demás factores, el agua siempredifunde desde donde es pura a cualquier solución, independientemente del soluto. Y, a bajas concentraciones, hay poco o ningún efecto del soluto particular; todos tienen el mismo efecto en la difusión independientemente de su impacto en la concentración numérica del agua.Es la concentración del soluto NO la concentración de moléculas de agua que están dirigiendo la difusión: todas otros factores manteniéndose constantes, el agua siempre se difunde desde donde hay una menor concentración de soluto hasta donde hay una mayor concentración de soluto. De esto, se puede concluir que es la pureza del agua, no su concentración, lo que impulsa la difusión. Esto puede parecer una sutil diferencia pero en realidad refleja algunas características muy profundas relacionadas con las leyes de la termodinámica. La pureza se relaciona con la entropía y se sabe que la entropía “impulsa” procesos espontáneos. Además, nuestra 'imagen mental', es decir, modelo, de lo que causa la difusión, no funciona —las moléculas solventes NO van de donde hay más de ellas a donde hay menos de ellas, tienden a ir de donde son más puras a donde son menos puras.
El efecto de la presión sobre la difusión
Una segunda razón clave por la que la descripción/modelo que describe la difusión como ocurriendo de 'alta concentración a baja' es deficiente es que no considera los efectos de la presión. La presión es la razón más familiar por la que los fluidos se mueven (viento, flujo de agua en tuberías, flujo sanguíneo en animales) pero estos movimientos no son difusión, son algo llamado 'flujo másico', un movimiento que depende únicamente de las diferencias de presión, y un movimiento que ocurrirá siempre que haya diferencias de presión y una trayectoria abierta para el flujo de fluido. Pero cuando el flujo másico es imposible (porque no hay 'camino abierto') la presión también puede influir en el movimiento difusional: el agua se difundirá de áreas de alta presión a áreas de baja presión. Esto es especialmente importante para las células que poseen tanto una membrana celular como una pared celular. Si bien la membrana permite que haya diferentes purezas del disolvente (agua) dentro vs. fuera de la celda, la pared permite que haya diferentes presiones dentro vs fuera de la celda, y tanto la pureza como la presión son importantes para dictar la difusión del agua.
Combinando los efectos de la pureza y la presión
Siendo iguales todas las demás cosas, el agua se mueve por difusión de regiones de mayor presión a regiones de menor presión y también de regiones de alta pureza a regiones de baja pureza. Estos dos factores pueden 'equilibrarse' entre sí y es posible tener difusión de NO entre un área de baja pureza y alta presión conectada a un área de baja presión y alta pureza. Mirando esto de otra manera, si tienes agua de baja pureza confinada en un recipiente rígido (es decir, una celda con una pared celular) y se pone en agua pura, el agua se moverá hacia la celda, aumentando la presión en la celda. Eventualmente, se alcanzará presión donde no haya más difusión en la celda. En este punto, las diferencias de presión entre el interior y el exterior coinciden con las diferencias de pureza entre el interior y el exterior.
Desafortunadamente, no existe un modelo fácilmente conceptualizado para la difusión de wateras líquidas que haya para la difusión de gases y solutos. Un modelo riguroso de difusión del agua líquida requiere la aplicación de conceptos a partir de un parámetro termodinámico llamado potencial hídrico. La idea básica es relativamente simple: la ósmosis (la difusión del agua líquida) es un proceso espontáneo y cualquier proceso espontáneo debe dar como resultado una disminución en la cantidad de energía disponible para realizar el trabajo (la 'energía libre'). Generalmente, dos factores clave afectan la energía libre del agua (su potencial hídrico): la presión (que aumenta su potencial hídrico) y la presencia de solutos (que disminuye su potencial hídrico). El agua líquida se difunde desde áreas de alto potencial hídrico a áreas de bajo potencial hídrico al igual que el calor fluye de áreas cálidas a áreas frías.
Para todos los organismos, y en particular aquellos sin pared celular, la membrana celular y la actividad celular permiten que se desarrollen diferencias de concentración de solutos entre el interior y el exterior de la célula, dando como resultado diferencias en la pureza del agua entre el interior de la célula y el exterior. Cuando se desarrollan diferencias de pureza, el agua fluirá dentro o fuera por difusión. Mientras las diferencias de pureza sean pequeñas, este movimiento puede eliminar las diferencias de pureza al hacer que el interior sea menos puro (si el agua fluye hacia afuera) o más puro (si el agua fluye hacia adentro). Sin embargo, dicho movimiento del agua también provocará que la célula cambie de volumen y si la célula se hincha o se contrae demasiado puede causar daños irreparables a la membrana, destruyendo así el funcionamiento celular. Como consecuencia, los organismos sin pared celular deben vivir en áreas donde la pureza del agua es similar a la que se encuentra dentro de sus células o deben tener estructuras/mecanismos que disminuyan la difusión y/o tengan la capacidad de eliminar el agua que se difunde en (por ejemplo, la vacuola contráctil de Paramecio) o adquirir agua para reemplazar la perdida por difusión (generalmente esto se logra adquiriendo el agua 'salada' y eliminando las sales).

Para las células con una pared (por ejemplo, plantas y hongos), la pared permite una nueva 'solución' para vivir en áreas donde la célula está más concentrada en solutos que en el ambiente externo (esta es la situación normal para la mayoría de los hábitats no marinos: lagos y arroyos de agua dulce y hábitats terrestres donde se encuentran los organismos inmerso parcial o totalmente en suelo cuya agua es generalmente bastante pura, es decir, con pocos solutos). Para estos organismos, la pared rígida permite que la presión aumente a medida que el agua fluye hacia las células. Esta presión actúa para reducir la difusión interna del agua y eventualmente se alcanza un equilibrio dinámico donde la alta presión y la baja pureza dentro de la celda equilibran la menor presión y mayor pureza fuera de la celda. El agua se mueve (se difunde, es decir, se mueve por ósmosis) dentro y fuera a la misma velocidad.
Plantas y hongos utilizan 'sistemas osmóticos' de varias maneras
Estructuralmente
El agua puede ser utilizada como 'bloques de construcción' cuando está confinada en una estructura que no se expandirá. Las células vivas, con la combinación de una membrana celular y una pared celular, son estructuralmente fuertes y las plantas y hongos las utilizan para formar estructuras rígidas que pueden soportar las fuerzas gravitacionales y del viento (también se discute en el capítulo 3). La evidencia de la importancia estructural del agua proviene de la observación del marchitamiento: si las plantas se ven privadas de una fuente de agua para reemplazar la perdida por evaporación, pierden integridad estructural. Algunas plantas y hongos producen algunas estructuras (por ejemplo, árboles, hongos de soporte) que no colapsan cuando se les priva de agua, pero para muchas plantasel acceso al agua es esencial para 'ponerse de pie' porque es la presurización de las células lo que proporciona rigidez. El centro de esta capacidad es una pared celular que tiene una alta resistencia a la tracción y resiste la expansión y, en consecuencia, permite la presurización.
Crecimiento
El crecimiento celular ocurre cuando las presiones internas exceden la fuerza de la pared celular y así cede a la presión dentro de ella. Los órganos (filamentos fúngicos, raíces y brotes) crecen como resultado de la expansión de las células individuales y la presión interna no solo tiene que empujar la pared celular sino que también puede tener que empujar (comprimir) el suelo a su paso. Así, para las plantas (tanto raíces como a veces brotes) y hongos, el crecimiento puede requerir la producción de una cantidad significativa de fuerza. La fuerza para impulsar este crecimiento proviene de la difusión del agua (ósmosis) y se pueden crear fuerzas muy significativas como resultado del movimiento del agua por su gradiente de potencial hídrico. Presiones de 2-4 bares (= 0.2-0.4 MPa [megapascales] = 2-4 atmósferas de presión = 30-60 libras por pulgada cuadrada) son comunes y pueden ser bastante efectivas, como se pudo haber dado cuenta cualquiera que haya observado un diente de león subiendo por una acera.
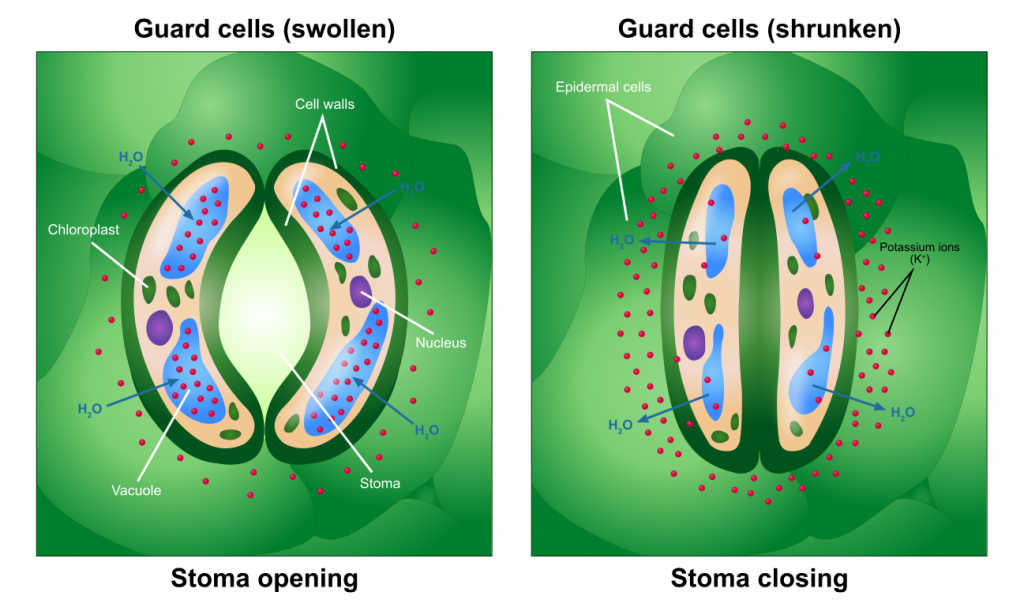
Células de guardia y estómagos
La apertura y cierre de estómagos se produce como consecuencia de los cambios en la presión de las células especializadas, células de guarda, que rodean el poro. La presurización de las celdas de protección, como resultado de la acumulación de soluto y la posterior difusión de agua en las celdas de protección, hace que las células se hinchen y formen una abertura (un estomato) entre ellas. Acrease en solutos en las células guardianasprovocará un movimiento del agua fuera de la célula, resultando en adrop en la presión y consecuentemente cierre estomático. Los desencadenantes que estimulan a las células guardianas a acumular o perder solutos han sido ampliamente estudiados e incluyen la luz y la concentración de dióxido de carbono. Los controles exactos pueden no ser los mismos para todas las especies. Al menos algunas plantas son capaces de regular los niveles internos de dióxido de carbono en un 'punto de fijación' que permite que la fotosíntesis proceda con poca inhibición debido a la falta de dióxido de carbono, al tiempo que minimiza la cantidad de agua perdida por la transpiración.
Movimiento de folleto
Similar a la acción de las células de guarda, varias plantas tienen hojas o folíolos que se mueven en respuesta a señales ambientales como la luz, el tacto y la sequía, dando como resultado hojas o folíolos cuya orientación varía según las circunstancias. Un ejemplo común son los 'movimientos del sueño' donde las hojas son horizontales durante el día y verticales por la noche. Estos movimientos son el resultado de cambios en la presión de las celdas 'pulvinares', ubicadas en puntos de pivote. Cambios relativamente pequeños en el tamaño de estas células se aprovechan como resultado de su ubicación y pueden causar cambios sustanciales en la posición de los órganos involucrados. (ver http://www.youtube.com/watch?v=U-PK13JEgk8 más abajo)
Ver

Un elemento de YouTube ha sido excluido de esta versión del texto. Puedes verlo en línea aquí: pb.libretexts.org/sdt34b/? p=302

Transporte de larga distancia
Floema
Ambos sistemas de transporte en plantas, el transporte de xilema y floema, operan como resultado de las diferencias de presión creadas en las 'tuberías' que se encuentran en estos tejidos. Las diferencias de presión se crean como resultado de la difusión del agua. En el floema, se crean presiones como resultado de la adición de solutos (azúcares) a las celdas de tubería, proceso conocido como carga de floema, que ocurre en regiones de la planta llamadas áreas 'fuente'. Como consecuencia de la carga del floema, el agua fluye hacia los elementos del tubo de tamiz del área fuente, y la presión aumenta, desencadenando un flujo en las tuberías. Las diferencias de presión y el flujo se mantienen porque los solutos no solo se cargan en las áreas de 'fuente', también se eliminan en las áreas de 'sumidero', las ubicaciones a las que se transportan los solutos; así se produce un gradiente de presión continuo en el floema, desde fuentes que proporcionan azúcares, hasta sumideros que consumen o almacenar azúcares. Las ubicaciones exactas que sirven como fuentes o sumideros pueden cambiar dependiendo de si una ubicación está produciendo azúcares (sacarosa) o consumiéndolos. El movimiento del floema puede ser hacia arriba de la planta (por ejemplo, desde sitios de almacenamiento en la raíz hasta meristemos apicales de brotes) o hacia abajo de la planta (por ejemplo, desde la fotosíntesis de hojas hasta el almacenamiento sitios en la raíz). La carga de sacarosa requiere energía metabólica ya que el ATP se usa para mover la sacarosa desde donde está menos concentrada a donde está más concentrada. Como fue el caso en las celdas de guarda, el movimiento del agua en los tubos de tamiz es pasivo una vez que se han agregado los solutos. Debido a que se trata de un flujo másico, no solo la sacarosa sino cualquier soluto que esté en el tubo del tamiz será transportado al fregadero. Los más comunes de estos otros solutos son los aminoácidos, pero otros compuestos que contienen nitrógeno se transportan en el floema junto con algunos elementos minerales (por ejemplo, K +, Mg 2 +, Ca 2 +)
Xilema

El transporte de agua en el xilema también es el resultado de diferencias de “presión”, pero en realidad son diferencias en la tensión más que diferencias en la presión. Mientras que la presión comprime los fluidos, la tensión separa los líquidos, así como tirar de una cuerda ejerce una fuerza que actúa para romper las moléculas de la cuerda. Sorprendentemente, en ciertas situaciones, el agua tiene una resistencia a la tracción sustancial y de hecho se puede tirar. Las paredes celulares de todas las células de una hoja se infunden y recubren con agua debido a la adhesión del agua a las paredes celulares y la cohesión del agua a sí misma. Cuando el agua deja una hoja por evaporación el agua restante se 'estira', se pone bajo tensión, debido a que el menor volumen restante de agua está cubriendo el mismo volumen original de celdas. Esta tensión se transmite al agua en las celdas conductoras del xilema y crea una diferencia de 'presión' (en realidad una diferencia de tensión) que puede elevar el agua por las celdas de tubería (no vivas). Lo que generalmente está impulsando el movimiento del agua por las tuberías (trach ei ds y vasos) es una tensión creada a medida que el agua se pierde debido a la transpiración. Sin embargo, el flujo de agua por el xilema también puede ocurrir incluso si hay poca transpiración mientras se produzca el crecimiento: la difusión del agua hacia las células en expansión puede crear una tensión para elevar el agua a las plantas en la primavera cuando las plantas carecen de hojas y las tasas de transpiración son muy bajas.
La pérdida de agua de la hoja es simple difusión: el vapor de agua a altas concentraciones (alta humedad del aire dentro de la hoja) se difunde a través de estomas abiertos hasta donde la humedad es menor fuera de la hoja. La humedad del aire dentro de la hoja se mantiene debido a que el agua líquida en las paredes celulares de las células mesófilas se evapora y reemplaza el agua que se ha perdido. Las tensiones generadas por la pérdida de agua hacen que las traqueides y los recipientes se compriman ligeramente ya que la presión exterior es 'normal' (una atmósfera) pero la presión en el interior es menor. Tenga en cuenta que esto contrasta con la situación en las células mesófilas vivas que están presurizadas porque tienen una membrana que les permite concentrar solutos. Si la tensión en la columna de agua se vuelve demasiado grande, la cavitación se produce a medida que se forman burbujas de aire cuando la columna de agua se rompe o cuando el agua se separa de los lados de la traqueidea o recipiente. En cualquier caso, la celda está 'cavitada' y ya no es útil para el transporte de agua.
Tanto en situaciones impulsadas por la transpiración como impulsadas por el crecimiento, y en el transporte de floema, hay un movimiento difusional de corta distancia que crea una diferencia de presión que puede resultar en un movimiento de larga distancia. Tanto en el xilema como en el floema el movimiento dentro de las tuberías NO es difusión, es un proceso mucho más familiar llamado flujo a granel. El flujo a granel es un medio mucho más efectivo para transportar materiales a largas distancias que la difusión, que solo es efectivo a distancias muy pequeñas (décimas de mm para líquidos). El flujo a granel está bloqueado por las membranas celulares y es impedido, pero no prevenido, por las paredes celulares. Las fosas que se encuentran en las paredes celulares de las traqueidas y los elementos tubulares del vaso proporcionan una trayectoria de resistencia relativamente baja para que el agua se mueva entre celdas adyacentes. El agua fluye aún más fácilmente a través de las placas de perforación de los vasos porque están completamente abiertas. En las células conductoras (vivas) del floema, los plasmodesmas conectan las células individuales y la savia del floema (que es esencialmente citoplasma que carece de orgánulos) se produce el flujo masivo de célula a célula a través de los plasmodesmas. El flujo a granel también es significativo en el suelo donde hay pasajes para que el agua fluya y donde tanto la gravedad como el 'tirón' por movimiento hacia las plantas pueden crear diferencias de presión.
Las presiones y tensiones encontradas en el tejido vascular reflejan estos mecanismos. Si se penetra un tubo de tamiz, la savia del floema fluye hacia afuera porque la presión dentro de la celda es mayor que la atmosférica, así como sangrarás si tu piel está cortada. En la mayoría de las circunstancias, si un traqueide o recipiente es penetrado, el agua NO fluye hacia afuera, sino que el aire fluye hacia adentro, reflejando el hecho de que el agua dentro del traqueide/recipiente estaba bajo tensión. De hecho, si se mide el volumen de una traqueidea/vaso a medida que se desarrollan las tensiones, disminuye ligeramente debido a la compresión desde el exterior. Debido a esto, los troncos de los árboles exhiben una disminución medible en la circunferencia durante el día a medida que aumentan la transpiración y las tensiones y rebotan durante la noche a medida que se alivian las tensiones y la planta se rehidrata en tiempos de poca o ninguna transpiración.
En raras ocasiones el agua en el xilema está presurizada.
Esta condición se describe como 'presión radical' y solo ocurre bajo ciertas circunstancias. La 'presión radical' es por el 'sangrado' (exudación) de un tallo decapitado. Bajo estas mismas condiciones especiales, si se perfora un vaso de xilema o traqueida individual también sangrará, a diferencia de la situación más normal descrita anteriormente. La presión radicular solo ocurre si el suelo está húmedo, las raíces están creciendo activamente y la transpiración es baja (por la noche o cuando no hay hojas presentes). En estas condiciones, los solutos (iones minerales) se acumulan en el xilema de la raíz debido a las acciones de las células radiculares, y porque la endodermis se comporta colectivamente como una membrana y es una barrera que evita que los solutos acumulados en el xilema radicular se escapen del tejido del xilema. De ahí que, como una célula vegetal individual viva que pueda presurizar porque la membrana celular permite acumular solutos, todo el xilema radicular puede acumular iones y presurizar. Este fenómeno es raro, tanto porque las raíces generalmente no son lo suficientemente activas como para acumular suficientes iones en el tejido del xilema y también porque un tirón desde la parte superior (creado por la transpiración o crecimiento) evita que se produzca una acumulación de presión.
Una situación final donde se presuriza el xilema es la que hace que la savia fluya (afuera) en los troncos de los arces a fines del invierno y principios de la primavera. 'Tapping', insertando un cilindro, en el xilema, da como resultado el sangrado de la savia que puede ser recolectada, concentrada y utilizada como fuente de azúcar. El flujo de savia de arce NO requiere actividad radicular, se puede observar en tallos retirados del sistema radicular. El flujo de savia de arce SÍ requiere ciclos de congelación/descongelación porque estos de alguna manera permiten que el xilema se presurice. Los azúcares que se encuentran en la savia del xilema provienen de los rayos de xilema cuyo almidón se convierte en sacarosa a fines del invierno. Por qué solo los arces, y algunas otras especies de árboles, exhiben este comportamiento generalmente se atribuye a aspectos de su anatomía de la madera. La mayoría de las especies arbóreas no se presurizan cuando se exponen a ciclos de congelación/descongelación.
Aunque el tejido del floema se penetra cuando se golpea el arce y aunque el tejido del floema se presuriza y transporta sacarosa, extremadamente poca de la savia recolectada de los arces se deriva del floema. Las células vivas (elementos de tubo de tamiz) del tejido del floema son capaces de taponar rápidamente los agujeros para evitar el 'sangrado' y la consiguiente pérdida de azúcar. Los arces que se golpean sangrarán del tejido del xilema durante varios meses, siempre y cuando estén expuestos a condiciones adecuadas y siempre que no haya un 'tirón' de la parte superior de la planta causado por el crecimiento o evaporación de las hojas. La savia de arce fluye cuando las hojas no están presentes y el crecimiento no ocurre. Si se pudiera aprovechar el tejido del floema se encontraría que tiene una concentración de azúcar mucho, mucho mayor (comparable al jarabe de arce) que la savia de arce, cuya dulzura apenas es detectada por los humanos hasta que se concentra.