
1.26: Interacciones que involucran condiciones
- Page ID
- 56436

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Un aspecto particularmente significativo e interesante de la vida orgánica es su interacción con su entorno circundante. Como indica el término, la interacción implica que ambos componentes se afectan entre sí: el ambiente afecta a los organismos y los organismos afectan al medio ambiente. El 'ambiente' incluye componentes que son vivos, es decir, otros organismos (el ambiente biótico), y componentes que no son vivos como rocas, nubes, agua, organismos muertos o partes de organismos muertos. El ambiente tiene propiedades o condiciones, como temperatura y concentración de oxígeno que comúnmente se consideran 'el ambiente abiótico' o el 'ambiente físico'. Las condiciones son las consecuencias de procesos físicos como radiación, difusión, convección; y estos procesos a veces están fuertemente influenciados por procesos bióticos (Figura 1), como la respiración (genera calor y dióxido de carbono, elimina oxígeno) y la fotosíntesis (absorbe luz, agrega oxígeno y agua, elimina el dióxido de carbono). Las condiciones presentes en un sitecán dictan si un organismo puede existir o no, y si existe, cómo se comporta. Esto a veces se describe como una interacción entre los organismos y el entorno físico, o abiótico.

Debido a que los organismos pueden afectar condiciones, pueden interactuar con otros organismos como resultado. Además de la interacción entre organismos mediada por condiciones, existen interacciones más directas entre organismos, como un organismo que se come a otro o un insecto que transporta polen de una planta a otra. Las interacciones son fundamentales para la disciplina de la ecología. De hecho, una definición de ecología es “un estudio de las interacciones entre los seres vivos y su entorno”.

Otra definición común de ecología es “un estudio de la distribución y abundancia de organismos” (Figura 2). La distribución y abundancia de organismos están controladas por las interacciones entre los organismos y su ambiente biótico y abiótico. Por ejemplo, la distribución de una planta podría estar limitada por la intensidad de la luz. La cantidad de luz disponible es consecuencia tanto del entorno físico (latitud, grado de nubosidad, ya sea que el sitio esté orientado al sur o al norte, etc.) como al ambiente biótico (presencia o ausencia de especies arbóreas para interceptar la luz). La distribución y abundancia de una planta en particular (especie 'A') también podría limitarse a la presencia o ausencia de un organismo que come la planta o tal vez por un organismo que poliniza la planta. En cualquiera de estos últimos casos, se podría considerar que los factores que determinan la distribución y abundancia de la segunda especie están controlando la distribución y abundancia de la especie A.
Otra definición de ecología es “un estudio de la organización biológica por encima del nivel individual”. Como hemos visto, los organismos tienen una estructura (forma, tamaño, organización de partes componentes), y también los organismos tienen funciones, hacen cosas como desarrollar, reproducir e intercambiar materia y energía con su entorno. De manera similar, los grupos de organismos (por ejemplo, poblaciones) tienen características estructurales y funcionales que pueden describirse y categorizarse, y se pueden buscar explicaciones sobre lo que determina su organización. Un punto importante es que la vida no se organiza simplemente a nivel de la célula y del nivel de un organismo; también hay una organización que involucra a grupos de organismos y este es el nivel de estudio para la disciplina de la ecología.

Dos entidades que estudian los ecologistas son poblaciones (grupos de individuos de la misma especie) y comunidades (conjuntos de especies en un área determinada). Sin duda, una población podría considerarse una 'cosa viva' y su distribución a través del paisaje es una manifestación de organización biológica y es una característica estructural de esta entidad. La abundancia de una especie (cuántos individuos están presentes por unidad del ambiente) es otra característica estructural de una población. El hecho de que las poblaciones tengan patrones de distribución y abundancia representa una organización, una estructura, y los procesos que determinan esta estructura son los mismos procesos discutidos anteriormente: interacciones entre organismos y sus entornos físicos y bióticos. Se pueden hacer argumentos similares respecto a la organización de las comunidades.
En nuestro examen del papel de las interacciones en la biología de los organismos, consideraremos en este capítulo las interacciones entre organismos y condiciones, examinando las condiciones clave que afectan a los hábitats terrestres y acuáticos, por qué estas afecciones afectan la función de un organismo y qué factores hacen que estas condiciones varíen, incluyendo cómo los propios organismos pueden controlar las condiciones. En el próximo capítulo, examinaremos interacciones más directas entre organismos, tratando de ver similitudes fundamentales en las formas en que los organismos interactúan entre sí. Finalmente, veremos la agricultura, una interacción de la que todos dependemos, y las interacciones (Capítulos 28-30) que son críticas para la agricultura.
Nuestro enfoque se centrará principalmente en los organismos, aunque muchos de los temas se superponen con la ecología y abordan los temas desde una escala más amplia.
TEMAS
- Temperatura
- Influencias de la temperatura
- Tolerancias de organismos
- Crecimiento del organismo
- Psicófilos y termófilos
- Maduración de frutos
- Temperatura como señal
- Influencia de la temperatura en otras condiciones
- Humedad
- Hábitats terrestres
- Hábitats acuáticos
- Salinidad
- Oxígeno
- Luz
- Corrientes
- Nutrientes
Temperatura
La temperatura es de importancia crítica para todos los organismos, afecta si pueden sobrevivir y lo que hacen. Incluso para los organismos que regulan su temperatura en un punto de ajuste, (es decir, homeotérmicas), la temperatura tiene consecuencias significativas. Pero la temperatura es aún más significativa para la gran mayoría de los organismos que no regulan su temperatura. Para tales organismos, denominados poiquilotermos, las condiciones térmicas de su entorno controlan su actividad metabólica. Aunque la temperatura es importante en todos los hábitats, es de relativamente menor importancia en la mayoría de los sistemas acuáticos debido a que las propiedades térmicas del agua amortiguan las fluctuaciones de temperatura. Pero los organismos en hábitats terrestres están sumergidos en un fluido (aire) que absorbe muy poca energía térmica y tiene una baja capacidad calorífica (es decir, poca capacidad de amortiguación). Esto hace que la temperatura de los sistemas terrestres sea mucho más dinámica en el tiempo y el espacio que en la mayoría de los sistemas acuáticos. Una de las razones clave por las que la temperatura es importante en los hábitats terrestres es porque afecta los niveles de evaporación y humedad. Si bien existen algunos hábitats terrestres donde la temperatura no es un determinante clave de las actividades, desde una perspectiva global la temperatura y la humedad son el factor principal que controla qué organismos están presentes y qué están haciendo.
Influencias de la temperatura en los organismos
Todos los organismos tienen un rango de temperaturas en el que pueden llevar a cabo las funciones que los definen y sostienen. Las temperaturas por encima y por debajo de este rango alteran su estructura y funcionamiento de varias maneras. A continuación se enumeran las alteraciones más importantes.
Membranas y temperatura

Las membranas son esenciales para la vida, regulan el movimiento molecular y realizan una variedad de otras funciones. La naturaleza física de las membranas se encuentra entre un sólido y un líquido; pueden describirse como “cristales líquidos” porque parte de su naturaleza es rígida y ordenada mientras que otras partes son similares a fluidos (Figura 4, discutida también en el Capítulo 3). Esta dualidad de estructura es importante para su función; necesitan ser en parte rígidas porque gran parte de su funcionamiento depende de la organización de sus partes; si estas partes están fuera de lugar, se interrumpe el funcionamiento. Al mismo tiempo, las membranas necesitan poder cambiar de forma y los componentes de la membrana sí necesitan poder moverse lateralmente; esto solo es posible debido a la fluidez de la membrana. La temperatura interrumpe el equilibrio entre fluidez y rigidez: las altas temperaturas hacen que las membranas sean más fluidas y las bajas temperaturas las hacen más rígidas (cristalinas). Los cambios en cualquier dirección son perjudiciales. Las características de membrana de los organismos que se encuentran en hábitats de diferentes temperaturas son diferentes en formas predecibles que se relacionan con el mantenimiento de cierto grado de 'fluidez'. A temperaturas más altas, es más probable que los lípidos contengan cadenas más largas de hidrocarburos y estas cadenas tienen más probabilidades de estar más saturadas. Ambas características hacen que las membranas en las que se encuentran sean más “sólidas” a cualquier temperatura en particular. En la misma línea, se ha encontrado que algunos organismos que experimentan variación anual en la temperatura ajustan su química de membrana de manera que mantiene un grado constante de fluidez a pesar de los cambios de temperatura. Así, el rango de tolerancia a la temperatura está determinado en parte por la naturaleza química de las membranas de un organismo.
Reacciones químicas coordinadas y temperatura
Un segundo factor involucrado en las interrupciones de la función del organismo tanto a altas como bajas temperaturas es el equilibrio de las innumerables reacciones químicas que se están produciendo dentro de las células. La velocidad de casi todas las reacciones químicas está fuertemente influenciada por la temperatura, con las velocidades que suben a medida que sube la temperatura. La mayoría de estas reacciones están en algún tipo de equilibrio para que, en general, no haya acumulación ni agotamiento de metabolitos. Este equilibrio puede alterarse tanto a temperaturas altas como bajas porque la sensibilidad a la temperatura no es la misma para todas las reacciones, por lo que las reacciones que están “en equilibrio” a algunas temperaturas pueden no estar a temperaturas más altas o más bajas. La bioquímica de las células es tal que el proceso de control (por ejemplo, bucles de retroalimentación) puede operar para lograr el equilibrio en las vías metabólicas, pero hay límites para estos procesos de control, y al menos algunos de los problemas con temperaturas altas y bajas pueden atribuirse a problemas con reacciones que se vuelven 'desequilibradas'.
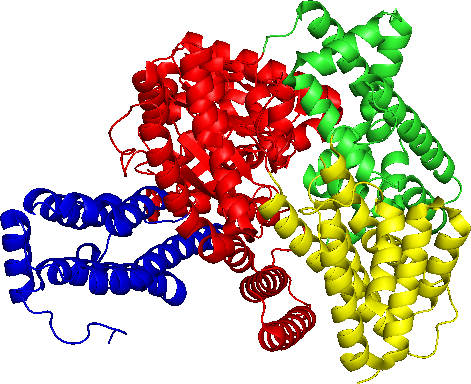
Enzimas y temperatura
El funcionamiento enzimático depende de una estructura tridimensional. Esta estructura puede ser alterada, un proceso llamado desnaturalización, por altas temperaturas debido a que el movimiento térmico es lo suficientemente significativo como para romper la unión relativamente débil que da cuenta de ciertos aspectos de la estructura enzimática. La temperatura a la que ocurre la desnaturalización varía con las proteínas pero está en el rango donde las temperaturas se vuelven letales para la mayoría de los organismos (25-40 C).
Problemas con la congelación
No es sorprendente que la congelación cause una variedad de problemas para los seres vivos. La expansión asociada con la congelación puede estallar las células, destruyendo membranas y paredes en el proceso. Para las plantas y hongos, que tienen un volumen sustancial de su estructura (el apoplasto) que se encuentra fuera de las membranas, la congelación (especialmente si ocurre lentamente) generalmente ocurre primero fuera del citosol, es decir, en el apoplasto, debido a su concentración de soluto sustancialmente menor. La congelación exterior provoca la difusión del agua desde el interior hacia el exterior, resultando en daños por desecación en el interior. Debido a estos efectos, un amplio grupo de plantas y hongos tienen su límite térmico inferior a 0 C, o ligeramente por debajo de él.
A pesar de esto, las plantas y los hongos sí viven en hábitats donde se produce la congelación. Para algunos, la supervivencia es el resultado de una parte invernada que se encuentra bajo tierra donde las temperaturas son más moderadas y nunca pasan por debajo del punto de congelación. Para otros, por ejemplo, árboles y arbustos, la existencia de partes perennes sobre el suelo refleja la capacidad de soportar períodos prolongados de temperaturas bajo cero. Los tejidos/células pueden evitar la congelación a través de dos mecanismos, uno que implica depresión del punto de congelación y el otro que implica sobreenfriamiento. La depresión del punto de congelación es una de las cuatro 'propiedades colligativas de las soluciones', cambios en las propiedades de una solución que ocurren con la adición de solutos, independientemente de qué soluto se agregue. Aunque la presencia de solutos en el citosol sí baja el punto de congelación, el efecto es relativamente pequeño (hasta -2 C, a las concentraciones de solutos que se encuentran típicamente en el citosol), y la protección contra la congelación por este mecanismo no es significativa para la mayoría de los organismos. El sobreenfriamiento puede resultar en caídas mucho más sustanciales en la temperatura a la que se produce la congelación. El sobreenfriamiento describe una situación en la que el agua líquida existe a temperaturas en las que generalmente se congela. Aunque el sobreenfriamiento puede ocurrir en agua pura, o casi pura, especialmente si no hay sitios para la nucleación del hielo (por ejemplo, bajo ciertas condiciones atmosféricas que causan gotas de lluvia sobreenfriadas), en los sistemas vivos el sobreenfriamiento parece ser una consecuencia de solutos, proteínas o glicoproteínas anticongelantes específicos, que de alguna manera impiden la formación de cristales. Dichos compuestos se encuentran no sólo en las plantas sino también en peces e insectos. El límite inferior de sobreenfriamiento es de alrededor de -40 C (que también es -40 F!!!!) , pero para muchas especies el límite de sobreenfriamiento está muy por encima de éste, en el rango de -10 C.
Un problema adicional de congelación, significativo para las plantas vasculares, es que cuando el agua se congela el aire disuelto se excluye del hielo, creando burbujas de aire en el hielo. Al descongelar estas burbujas permanecen. Esto es potencialmente un problema para el transporte de xilema porque las tensiones que se desarrollan durante el transporte del xilema provocarán la expansión de las burbujas y cavitación (bloqueo de aire) de los vasos y traqueides, interrumpiendo el transporte de agua del xilema. Este problema puede evitarse mediante la producción de nuevas celdas conductoras de agua a principios de la primavera, por la presurización del xilema ('presión radical') que ocurre en al menos algunas especies como resultado de la absorción de solutos en la primavera, o como resultado de procesos físicos que pueden eliminar las burbujas.
Tolerancias
Todos los organismos tienen límites térmicos altos y bajos; si un organismo alcanza esa temperatura muere. La exposición crónica a temperaturas más moderadas también puede ser letal, de una manera relacionada con la duración de la exposición. Algunas plantas serán matadas por una exposición a corto plazo a 38 C, mientras que 36 C es letal si la exposición es superior a 60 minutos.
Tolerancias extremas

La mayoría de los organismos tienen tolerancias a altas temperaturas de alrededor de 40 C y tolerancias a bajas temperaturas de 0 C, pero hay etapas de muchos organismos que tienen un rango de tolerancia mucho más amplio. Esta tolerancia a menudo se asocia con una etapa en el ciclo de vida del organismo que se dedica a la dispersión, por ejemplo, esporas, semillas. Estas estructuras sirven para perpetuar el organismo a través de un tiempo de condiciones desfavorables. Junto con la tolerancia a temperaturas extremas, normalmente viene una tolerancia a la desecación y a una serie de 'insultos' que normalmente matarían las células (por ejemplo, radiación ultravioleta, pH extremo, falta de oxígeno). Esta tolerancia es consecuencia de una estructura celular alterada que simultáneamente aumenta la tolerancia y disminuye la actividad metabólica. En definitiva, la estructura de la (s) célula (s) se vuelve cada vez más inanimada y cada vez menos afectada por condiciones extremas. Por lo general, este estado de 'animación suspendida' implica varios o todos los siguientes: acumulación de materiales como almidón o aceites que sirven como reservas de energía y carbono reducido, cambios en la estructura de la membrana, cambios en la estructura de las proteínas, cambios en orgánulos (en eucariotas), reducción en citoplasmáticos volumen, desecación y, para ciertas células, un engrosamiento de la pared celular (o a veces la adición de una pared a una célula que anteriormente carecía de una).
Generalmente, estas estructuras tolerantes son células individuales pero en algunos organismos los tejidos o todos los organismos experimentan cambios comparables en estructura y función. En el reino vegetal, tales tejidos tolerantes suelen estar presentes en las semillas y a veces en los meristemos apicales. Dicha tolerancia se puede encontrar ocasionalmente en otras estructuras y también se puede encontrar en plantas enteras, especialmente las que se encuentran en ambientes congelados o desiertos. La siguiente tabla enumera estructuras procariotas, protistas, fúngicas y vegetales que son particularmente tolerantes a condiciones extremas. Si bien la tabla solo tabula tolerancias a bajas temperaturas, es importante darse cuenta de que, en general, la tolerancia a altas temperaturas, así como la tolerancia a la desecación, se correlaciona con la tolerancia a bajas temperaturas. Por ejemplo, la mayoría de las semillas toleran fácilmente la exposición prolongada a condiciones congeladas; también toleran la desecación y la exposición a altas temperaturas (por ejemplo, 50 C) que normalmente matarían las plantas.
|
grupo |
estructura |
notas |
|
algunas bacterias |
endosporas |
la mayoría son muy resistentes a altas y bajas temperaturas y sobreviven por períodos prolongados |
|
la mayoría de las bacterias |
quiste microbiano (=exosporas) |
las células bacterianas aumentan el grosor de la pared celular y contraen el volumen del citoplasma |
|
cianobacterias |
akinete |
un akinete es una espora especializada que se encuentra sólo en algunas cianobacterias |
|
euglenoides |
quiste |
formación de quistes a menudo se desencadena por cambios en los niveles |
|
dinoflagelados |
quiste |
los quistes no tienen flagelos y producen una pared celular con celulosa, características que normalmente NO están presentes en los dinoflagelados |
|
diatomeas |
células en reposo y esporas |
las esporas tienen paredes de sílice engrosadas, las células en reposo no; estas estructuras pueden tener requisitos para la germinación. Las esporas de formas marinas pueden ser componentes importantes del registro fósil |
|
cigomicetos |
esporas, cigoesporas |
las cigoesporas que están asociadas con el sexo y son multinucleadas tienen una pared celular mucho más gruesa y son más tolerantes al abuso que las células normales |
|
basidiomicetos |
esporas tanto asexuales como sexuales |
además de esporas, algunas formas producen esclerocios, hifas desecadas y modificadas que son inactivas y tolerantes a los extremos |
|
ascomicetos |
esporas tanto asexuales como sexuales |
(ver descripción anterior para basidiomicetos) |
|
musgos |
plantas enteras, esporas |
las esporas de la mayoría de las especies son tolerantes a una variedad de tratamientos; además, toda la planta gametofita de muchos musgos tolera tanto las temperaturas de congelación como la desecación |
|
helechos y otras plantas vasculares sin semillas |
esporas, raramente plantas enteras |
a diferencia de los musgos y como la mayoría de las plantas de semillas, los helechos generalmente no pueden tolerar la desecación, aunque las esporas de helechos toleran este y otros tratamientos extremos. Solo unas pocas especies tienen partes sobre el suelo que hibernan en condiciones de subcongelación. |
|
coníferas |
granos de polen, semillas, plantas enteras |
como grupo, muchas coníferas son más tolerantes a la congelación que la mayoría de las plantas con flores, evidenciado por la presencia de coníferas a altas latitudes y elevaciones; sin embargo, las esporas NO son especialmente tolerantes; generalmente solo ocurren en tejidos hidratados; |
|
cícadas |
semillas solo para la mayoría de las especies |
solo unos pocos pueden tolerar temperaturas bajo cero, la mayoría están restringidas a áreas más cálidas |
|
gnetófitos |
solo semillas |
solo efedra tolera temperaturas bajo cero |
|
ginkgo |
polen, semillas, partes sobre el suelo |
el árbol es tolerante a la zona 3 del USDA, que tiene temperaturas de hasta -40 |
|
plantas con flores |
polen, semillas, partes elevadas de algunas especies |
bastantes plantas con flores se han adaptado a temperaturas muy por debajo del punto de congelación; también hay una serie de especies que pueden tolerar temperaturas superiores a 40 C |
Influencias de la temperatura en el crecimiento
Debido a que la temperatura afecta la velocidad de las reacciones químicas y porque las reacciones químicas determinan lo que hace un organismo, y en particular la velocidad a la que se hacen las cosas, la temperatura tiene un profundo efecto en el funcionamiento del organismo. Los efectos combinados de la actividad química de un organismo se denominan metabolismo y las tasas metabólicas comúnmente se duplican a triplicar con un aumento de temperatura de 10 C (18 F). Si bien la mayoría de los estudiantes generalmente asumen que la oportunidad de 'hacer más' como consecuencia de una mayor tasa metabólica es algo deseable, este no es siempre el caso. Las tasas metabólicas más altas requieren más alimentos porque el metabolismo de un organismo corre sobre la respiración celular. Hasta cierto punto, las poiquilotermas sobreviven a periodos de bajas temperaturas porque cuesta muy poco mantenerlas bajo estas condiciones. Las tasas metabólicas más bajas significan menores necesidades de oxígeno; en consecuencia, muchas plantas pueden sobrevivir a condiciones bajas de oxígeno (típicamente provocadas por inundaciones) mucho más fácilmente si las temperaturas son bajas.
Crecimiento y grado de crecimiento días Para cualquier organismo una de sus actividades más significativas es el crecimiento, la adquisición de materiales, y la posterior utilización de materia y energía para hacer que el organismo sea más grande y, en última instancia, acoplado a la producción de nuevos organismos. El efecto de la temperatura sobre el crecimiento se ve muy bien en el concepto de días de grado de crecimiento, una estadística que integra tiempo y temperatura y se utiliza para predecir el progreso de una amplia variedad de cultivos durante la temporada. Si bien los detalles varían entre cultivos, e incluso entre diferentes variedades de un cultivo específico, la idea básica es que se pueda predecir la etapa de crecimiento de una especie de cultivo haciendo un seguimiento del número de días que el cultivo ha pasado a diferentes temperaturas. Por ejemplo, si las temperaturas diarias tuvieran un máximo de 86 F y un mínimo de 70 F, el maíz podría tardar 100 días en alcanzar la madurez (es decir, el tiempo de cosecha); si las temperaturas fueran más frías, con un máximo diario de 80 F y el bajo de 60 F, podría tomar 140 días. Los agrónomos han desarrollado modelos para predecir cuánto tiempo tardará el cultivo en alcanzar la madurez en base a la acumulación de algo llamado 'día de grado de crecimiento'. Con base en su temperatura, a cada día se le asigna un cierto número de días de grado de crecimiento (GDD's), con días cálidos ganando más que los fríos. Una fórmula típica que calcula los días de grado de crecimiento es la siguiente:
GDD = [(Tmax + Tmin) /2] — 50
Dados los días enumerados anteriormente, el día más cálido vale (86+70) /2—50 = 28 GDD, y el día más frío vale (80+60) — 50 = 20 días. Si el maíz necesita 2800 días GDD para alcanzar la madurez lo hará en 2800/28 = 100 días a las temperaturas más cálidas y 2800/20 = 140 días a las temperaturas ligeramente más frías.
Este es un ejemplo de un 'modelo empírico', uno que intenta predecir cosas a partir de observaciones pero no necesariamente basado en una comprensión de cómo opera un sistema. La base fundamental para el modelo es la observación de que los cultivos crecen más rápido cuando es más cálido. El modelo es exitoso a pesar de que tiene un enfoque bastante crudo de la relación entre crecimiento y temperatura. Hay una serie de características del modelo que son útiles para apreciar:
- un GDD representa una unidad de crecimiento y la idea básica es que el desarrollo vegetal puede representarse como una acumulación de estas unidades de crecimiento.
- un GDD también representa el producto del tiempo y la temperatura; aunque esto se representa en sus unidades ('grados-días'), no es obvio en la ecuación anterior porque el factor tiempo es siempre de 1 día y no está incluido; la ecuación podría escribirse:
- GDD = [(Tmax + Tmin) /2 -50] grados * [1.0] día
- el término temperatura tiene dos componentes, el primero representa un tipo de promedio, una representación de una variable dinámica (cambiando con el tiempo), en este caso, la temperatura. Este promedio particular (sumar los valores extremos y dividir por 2) es conveniente, ya que es comúnmente utilizado por los meteorólogos para reflejar la temperatura diaria. A pesar de que es un tipo crudo de promedio, basado únicamente en dos valores, funciona bien. El otro término de temperatura (50 en las ecuaciones anteriores) podría describirse como una 'temperatura base', la temperatura mínima a la que no se produce crecimiento alguno. Aunque la ecuación potencialmente predice un 'crecimiento negativo' imposible, si la temperatura promedio está por debajo de la temperatura base, esta predicción rara vez es el caso porque en la mayoría de las situaciones los cultivos no se siembran hasta después de que la temperatura promedio la temperatura diaria excede la temperatura base.
- el concepto GDD ha sido utilizado con éxito para una amplia variedad de cultivos y también con insectos, generalmente con modificaciones de la 'temperatura base' y/o cambios en el término de temperatura promedio (por ejemplo, dejando que la temperatura máxima nunca supere 86)
Si se intentara idear un 'modelo mecanicista', uno que opere con base en los mecanismos de crecimiento, sería mucho más desafiante. El crecimiento es un proceso complejo que es consecuencia de una variedad de reacciones químicas que ocurren simultáneamente. La mayoría de estos están claramente afectados por la temperatura, pero predecir el efecto neto de la temperatura en todos ellos sería particularmente desafiante. Ciertamente, los procesos asociados al crecimiento están controlados por la temperatura, pero las interrelaciones asociadas con el crecimiento serían difíciles de dilucidar.
Variación en los rangos de temperatura para el crecimiento Si bien la gran mayoría de hongos, plantas y protistas tienen una temperatura óptima para el crecimiento en el rango de 25-35 C, existen algunas excepciones. Varias plantas tienen tasas máximas de crecimiento muy por debajo de 35 C. No es sorprendente que estas plantas crezcan en hábitats más fríos. Si bien existen varias razones posibles para ello, incluida la posibilidad de que estos organismos tengan membranas que funcionen de manera más apropiada a temperaturas más bajas (ver arriba), o la interacción entre temperatura y humedad (ver más abajo), otra explicación involucra la sensibilidad a la temperatura de fotosíntesis y respiración. Como se mencionó anteriormente, la respiración está estrechamente ligada a la temperatura y para la mayoría de las plantas alcanza un máximo en el rango de 30-35 C. La respuesta de la fotosíntesis es diferente: generalmente es menos sensible a los cambios de temperatura y, para las plantas C3, generalmente alcanza un máximo a temperaturas inferiores a 30 C, a veces muy por debajo (Figura 7). La razón por la que la fotosíntesis es menos sensible a la temperatura se debe en parte al hecho de que parte de la química clave es fotoquímica y las reacciones fotoquímicas no dependen fuertemente de la temperatura. Un factor adicional es que el dióxido de carbono, un reactivo en la fotosíntesis, debe disolverse en agua para llegar al sitio de la fotosíntesis (el cloroplasto) y la solubilidad del dióxido de carbono disminuye a medida que aumenta la temperatura. Debido a esto, las plantas C4, que tienen mecanismos para concentrar el dióxido de carbono, generalmente tienen temperaturas más altas para la fotosíntesis máxima que las plantas C3.

Al considerar los efectos de la temperatura sobre la fotosíntesis, la respiración y el crecimiento, se puede considerar la fotosíntesis como 'hacer alimentos' y la respiración como 'comer alimento'. A medida que la temperatura aumenta en el rango de 25-35°C, el apetito de las plantas continúa aumentando rápidamente mientras que la tasa de producción de alimentos disminuye o disminuye. El efecto neto de esto es que a temperaturas más altas cada vez hay menos alimentos para impulsar el nuevo crecimiento. En consecuencia, el crecimiento de las plantas a menudo disminuye a temperaturas más bajas de lo que cabría esperar.
Psicófilos, Criófilos y Termófilos

Varios organismos, en particular ciertas bacterias pero también algunos hongos y protistas, se denominan 'psicrófilos' o 'criófilos' porque lo hacen mejor a bajas temperaturas, a veces a temperaturas por debajo del punto de congelación. Si bien se mantiene el patrón básico de aumento de actividad con aumento de temperatura, el rango de actividad se desplaza a temperaturas mucho más bajas: 0 a 10°C y a veces -10 a 0°C Estos organismos tienen membranas modificadas y, para los que operan por debajo del punto de congelación, compuestos anticongelantes que les permiten operar a temperaturas tan bajas. Dado que un mecanismo muy común de conservación de alimentos son las temperaturas bajas/congelantes, estos organismos pueden plantear problemas para la industria alimentaria. Un hongo criófilo ha resultado ser el culpable de la 'enfermedad de nariz blanca' de los murciélagos, una enfermedad que recientemente ha diezmado poblaciones de murciélagos que hibernan en cuevas con temperaturas en el rango de 5-15°C. El comportamiento del murciélago de bajar la temperatura corporal durante el invierno (con el fin de ahorrar energía) ha proporcionado a estos hongos condiciones perfectas para su crecimiento.
En el otro extremo del rango de tolerancia se encuentran los termófilos, que operan muy por encima del rango de actividad normal de 0-35°C Al igual que con los psicrófilos, la gran mayoría de los termófilos son procariotas, la mayoría de las veces arqueas, pero hay algunos eucariotas termófilos, todos ellos hongos. Varios de estos hongos termófilos son componentes importantes de las operaciones de compostaje a gran escala importantes para la producción de hongos comerciales donde la generación combinada de calor metabólico del compost puede elevar la temperatura del mantillo a 80°C.
Maduración de la fruta
Se mencionará un proceso adicional afectado por la temperatura por su importancia comercial: la maduración de los frutos. Además de la influencia de la temperatura en el crecimiento de los frutos, la temperatura continúa afectando a los frutos después de que cesa el crecimiento, en el proceso de desarrollo describimos como maduración. Este proceso trae consigo importantes cambios en las características (aroma, color, sabor, textura) que son de importancia tanto para los consumidores como para los productores. Este proceso de desarrollo a menudo se controla de diversas maneras por la temperatura, y la temperatura puede tener un impacto significativo en el valor comercial de una variedad de cultivos frutales al influir en la aparición de rasgos deseables e indeseables. En un sentido más general, la temperatura puede influir en las características ecológicamente significativas de los frutos que afectan su papel en la dispersión de semillas al influir en su atractivo para los frugívoros.
Temperatura como Cue

Para muchas plantas, y algunos hongos y protistas, la temperatura proporciona una señal importante que se utiliza para coordinar el crecimiento y otras actividades con cambios estacionales en las condiciones. Así como la gravedad organiza la actividad de la planta en el espacio, la temperatura puede coordinar la actividad de la planta a tiempo. En esta situación, la temperatura no es solo una condición a la que responde el organismo, es una señal que transmite información sobre cómo serán las condiciones en el futuro, similar a la forma en que la duración del día (fotoperíodo) puede proporcionar información sobre las próximas temporadas. Sin embargo, dado que el ruido en la señal de temperatura es sustancial, usarlo para predecir condiciones futuras es más complicado: un retorno a las condiciones cálidas después de las frías podría ser un deshielo de enero o podría ser algo real (primaveral). Para muchas plantas, la primavera se percibe como un período cálido después de un período medido de frío. Un ejemplo de esto se ve en los patrones de germinación de muchas semillas. Las semillas de la mayoría de las plantas son shedin estado latente. Aunque algunas semillas no tienen requisitos específicos de germinación, simplemente necesitan unas semanas y son capaces de germinar (esto ha sido seleccionado en muchas especies de cultivos), muchas semillas requieren condiciones específicas o una serie de condiciones específicas para poder germinar. Especialmente en hábitats templados, a menudo es un tratamiento de temperatura fría que 'romperá' la latencia, es decir, permitirá que la planta en la semilla reanude el crecimiento (Figura 9). Este patrón es beneficioso porque evitaría que las semillas germinaran y con ello se volvieran susceptibles al frío hasta después de las duras condiciones del invierno. Debido a que las prácticas hortícolas originales desarrolladas para romper la latencia de las semillas implicaban estratificar las semillas y mantenerlas húmedas y frescas, dicho tratamiento se denomina “estratificación”. La parte clave del tratamiento es el tratamiento de temperatura, no la estratificación. Contrario al patrón metabólico normal, donde la actividad es promovida por temperaturas más cálidas (es decir, temperaturas más cálidas producen más respuesta que las más bajas), en la estratificación la actividad de respuesta es promovida por temperaturas más bajas, teniendo temperaturas más frías mayores efectos que las superiores. Típicamente las temperaturas deben estar por debajo de 50°F (10°C) para ser efectivas y ser más efectivas a medida que la temperatura disminuye hasta 0°C. Las temperaturas por debajo de 0°C, y los tratamientos de temperatura cuando la semilla se seca, no son efectivos, lo que indica que la actividad metabólica es esencial para la respuesta. El proceso se puede modelar de manera similar a los días de grado de crecimiento, excepto en este caso lo que se acumula, denominado “unidades de enfriamiento”, requiere temperaturas por debajo de cierto umbral (típicamente 10°C), y se acumulan más unidades de enfriamiento a medida que la temperatura disminuye a 0°C. acumula suficientes unidades de frío su latencia se rompe; en este punto, el comportamiento del embrión es típico de la mayoría de los organismos, y la actividad (crecimiento) es promovida por temperaturas más cálidas.
Además de romper la latencia de los embriones en las semillas, los tratamientos con frío también son una señal muy importante para romper la latencia de los meristemos apicales de brotes (cogollos). Los jardineros caseros a menudo traen brotes de árboles y arbustos en flor (por ejemplo, manzana) al interior para 'obligarlos' a florecer temprano (Figura 10). Esta práctica funciona siempre y cuando el brote haya estado expuesto a temperaturas más frías durante el tiempo suficiente antes de calentarlo. Los brotes a menudo no forzarán en diciembre sino que lo harán en febrero. La cantidad exacta de 'tiempo de enfriado' que necesita una toma antes de que responda a temperaturas más cálidas varía. Este fenómeno es a veces la razón por la que ciertos árboles pueden no ser capaces de cultivarse en latitudes del sur: nunca reciben temperaturas suficientemente frías como para hacer que emerjan de la latencia.

La acumulación de unidades de frío también es una señal para que algunas plantas florezcan, un proceso denominado 'vernalización. 'En algunas situaciones el tratamiento en frío en sí es el detonante de la floración; en otras situaciones, el tratamiento en frío simplemente activa un interruptor y permite que la planta florezca en respuesta a una segunda señal (por ejemplo, fotoperíodo) que anteriormente no provocaría ninguna respuesta.
Efecto de la temperatura en otras condiciones

La temperatura también es importante porque interactúa con una serie de otras condiciones. En hábitats terrestres, probablemente la interacción más significativa sea entre temperatura y humedad. La temperatura afecta las tasas de evaporación, y ocasionalmente condensación. La evaporación ocurre cuando una molécula de agua individual se mueve lo suficientemente rápido como para escapar de las fuerzas cohesivas de sus vecinos. Dado que la velocidad de las moléculas es una función de la temperatura, cuanto más cálida es, más probable es que se produzca la evaporación. La tendencia de una sustancia a evaporarse se refleja en una propiedad llamada presión de vapor, que mide la cantidad de la sustancia (en este caso agua) presente en el estado de vapor cuando el líquido y el aire están en equilibrio. La Figura 11 muestra el efecto muy fuerte que tiene la temperatura sobre la presión de vapor. Bajo ciertas circunstancias, la evaporación estará directamente relacionada con la presión de vapor y en consecuencia se duplicaría aproximadamente con cada aumento de temperatura de 10 o C. En ambientes donde el agua perdida es difícil de reemplazar, la temperatura puede ser de gran importancia para las tasas de desecación y la supervivencia de los organismos. Para plantas y hongos, los factores combinados de temperatura y disponibilidad de agua son de suma importancia para determinar la abundancia y actividad de las especies. Tenga en cuenta que aunque la temperatura tiene una influencia importante en la evaporación por su efecto sobre la presión de vapor, también son importantes otros factores como la humedad del aire, la superficie de contacto entre el aire y el agua, y el grado de mezcla (convección) del aire por encima de la superficie hidratada. Finalmente, aunque la temperatura afecta la evaporación, la evaporación, a su vez, afecta la temperatura. En el caso de las hojas de las plantas, la evaporación puede hacer que las hojas sean significativamente (más de 2 o C) más frías que la temperatura del aire.
La temperatura también afecta la disponibilidad de ciertos compuestos al afectar su solubilidad en agua. Si bien la solubilidad de muchos sólidos en el agua aumenta con temperaturas más altas, lo contrario es cierto para los gases, y en particular para el dióxido de carbono y el oxígeno. Esto es particularmente significativo para la fotosíntesis y, como se describió anteriormente, es parte de la razón por la cual la respuesta de la fotosíntesis con el aumento de la temperatura no es comparable a la de la respiración aunque ambas implican una gran cantidad de reacciones químicas mediadas por enzimas que generalmente se potencian a mayor temperaturas.
Otra situación en la que la disminución de la solubilidad de los gases a altas temperaturas puede ser un problema son los suelos inundados. El efecto de las inundaciones es más dañino a altas temperaturas que a bajas. Una variedad de factores contribuyen a esto: (1) disminución de la solubilidad del oxígeno a temperaturas más altas, (2) aumento de las tasas de respiración del suelo a temperaturas más altas como lo lleva a cabo la suma de organismos presentes en el suelo (raíces de plantas, hongos, bacterias, protozoos, etc.); disminución de los medios respiratorios incrementados oxígeno (3) aumentó los requerimientos de oxígeno para las raíces de las plantas debido a las mayores tasas metabólicas asociadas a temperaturas más altas.
Agua y hábitats terrestres
Junto con la temperatura, la humedad es una variable ambiental clave que dicta la distribución de organismos en hábitats terrestres. Las razones de esto deben ser claras:
- el agua es un componente esencial de los tejidos vivos y un reactivo en muchas reacciones esenciales, incluyendo la fotosíntesis y la hidrólisis de ATP
- en hábitats terrestres, el agua casi siempre se pierde a la atmósfera en el proceso de evaporación
- para los autótrofos terrestres, que obtienen dióxido de carbono de la atmósfera, la evaporación es aún más probable que ocurra ya que los organismos deben exponerse a la atmósfera para adquirir carbono
- para plantas y animales, el agua es el medio en el que los materiales se mueven cuando se transportan y para los animales, el agua es la base para la excreción
Tolerancia al secado
La gran mayoría de los organismos mantienen sus condiciones de humedad a un nivel particular a través del proceso familiar de homeostasis. La mayoría de las plantas (y la mayoría de los organismos en general) serían consideradas homiohídricas, es decir, mantienen sus niveles de agua en un “punto de ajuste”. Para ello se requiere que un organismo ajuste ya sea la ganancia de agua o la pérdida de agua. En cierta medida, las plantas ajustan la ganancia de agua: cuando se secan, la fuerza impulsora para la absorción de agua aumenta y esto puede resultar en un flujo incrementado (¡recuerda la ecuación del flujo!). Sin embargo, las plantas solo tienen una capacidad bastante limitada para “secarse”, y cuando comienzan a desecarse, sus ajustes más significativos implican reducir la pérdida de agua en lugar de aumentar la ganancia de agua. Cuando las plantas experimentan déficits de agua, su respuesta es reducir la pérdida de agua al: (1) reducir la permeabilidad de la planta al agua cerrando los estomas, y (2) reduciendo la superficie para la pérdida, generalmente desprendiendo hojas. Nuevamente, recuerde la ecuación de flujo y cómo estos cambios se relacionan con ella. La pérdida de hojas es especialmente obvia en hábitats estacionalmente secos donde las especies leñosas pierden todas sus hojas y las especies herbáceas pasan la estación seca como bulbos u otras partes subterráneas, ambos grupos producen y poseen hojas solo cuando las condiciones son más mésicas.
Estas medidas para reducir la pérdida de agua vienen al precio de reducir la capacidad de una planta para alimentarse por sí misma. El efecto muy significativo que la humedad tiene en las distribuciones de las plantas refleja el hecho de que el éxito individual, poblacional y de especies implica equilibrar la pérdida de agua y la ganancia de carbono. En algunas situaciones, una especie puede tener éxito al poder adquirir 'su propia' fuente de agua. Las raíces de alfalfa y mezquite a menudo penetran lo suficientemente profundo como para aprovechar fuentes de agua subterránea no disponibles para otras plantas Sin embargo, la mayoría de las plantas de un área sharea fuente de agua común, el suelo, es decir, las raíces de muchas especies están ocupando el mismo volumen de suelo que otras especies, haciendo que el suministro de agua sea uniforme para la mayoría de las especies que viven en una zona. En consecuencia, la conservación por una especie o individuo solo deja aguaque puede ser tomada por otras especies. Las especies tienen éxito al manejar su crecimiento total y sus patrones de crecimiento. Esto representa un área de diversificación entre diferentes especies.
Los organismos homehídricos son intolerantes a la desecación y mueren si su estado hídrico cae por debajo de cierto nivel; la mayoría de las plantas no pueden recuperarse de una pérdida de 10-15% de su agua. Probablemente la razón principal de esto en las plantas vasculares es la “disfunción catastrófica del xilema” que es consecuencia de la cavitación y los bucles de retroalimentación positiva relacionados con la cavitación. Recordemos que los vasos individuales y las traqueidas pueden cavitar si el agua en ellos se tira con demasiada fuerza, resultado de la pérdida de agua en un momento en que existe una restricción de adquisición de agua a medida que el suelo se seca. La cavitación da como resultado una pérdida en una parte del sistema conductor del xilema y esto hace que la cavitación sea más probable en los elementos conductores restantes (ver la ecuación del flujo); mayor resistencia al flujo como resultado de la cavitación significa que tiene que haber una mayor tracción (mayores tensiones) para adquirir la misma cantidad de agua. Por lo tanto, hay un bucle de retroalimentación positiva: la cavitación hace más probable la cavitación, lo que hará más probable la cavitación. La disfunción catastrófica del xilema (sin capacidad para transportar agua) puede resultar rápidamente después del primer evento de cavitación, dejando a la planta sin capacidad de rehidratarse.
Hay un pequeño número de organismos incluyendo muy pocos animales (tardígrados), algunos protistas, y algunas plantas, que son poikilovídricos: su estado hídrico no está fuertemente regulado sino que se le permite asumir el nivel dictado por el ambiente en el que se encuentran. Obviamente, estos organismos solo pueden tener éxito (asumiendo que viven en un ambiente que se seca al menos parte del tiempo) si son tolerantes a la desecación, fenómeno que es muy raro en los organismos, aunque pueden producir partes como esporas y semillas que son tolerantes. Los organismos poiquilídricos pueden perder hasta el 90% de su agua, pero aún pueden revivir cuando el agua vuelve a estar disponible. El único grupo de plantas donde la tolerancia a la desecación es común es en plantas no vasculares, muchos musgos y algunas hepáticas, (irónicamente grupos que a menudo se consideran restringidos a ambientes húmedos); también está presente algunos helechos y musgos y en muy pocas plantas semilleras.


Si bien hay algunos musgos que están restringidos a hábitats húmedos, la mayoría son tolerantes a la desecación y muchos son particularmente prominentes en situaciones áridas, tanto en hábitats áridos (desiertos) como en porciones áridas de hábitats más mésicos, por ejemplo, que crecen en rocas que no tienen capacidad de almacenar agua. Algunas de estas especies tienen características para disminuir la pérdida de agua, por ejemplo, una respuesta de secado que implica el desprendimiento de 'hojas' alrededor del tallo, disminuyendo la superficie expuesta a la atmósfera. Sin embargo, muchos musgos son capaces de tolerar la desecación extrema y todo el organismo, no solo una parte selecta, y pueden permanecer viables después de someterse a ciclos repetidos de desecación. Están inactivos durante los periodos secos pero son capaces de reanudar rápidamente la actividad cuando se humedecen.


Tolerancia a inundaciones
En el otro extremo, los organismos terrestres pueden verse afectados por excesos de agua. Generalmente este es un efecto indirecto de la falta de oxígeno que se discutió anteriormente al considerar las interacciones entre temperatura y oxígeno.
Agua y Ambientes Acuáticos
Salinidad
Se podría suponer que no hay problemas asociados con el agua en un ambiente acuático ya que el agua es abundante. Sin embargo, el agua juega un papel importante en dictar los organismos presentes en cualquier ambiente acuático en particular, principalmente por el proceso de difusión. Debido a que el agua tiende a difundirse de regiones donde es más pura a regiones donde es menos pura, los seres vivos se ven fuertemente afectados por la pureza del agua en la que viven y esto está determinado principalmente por la ssalinidad del agua. Toda la vida tiene la capacidad tanto de acumular como de generar solutos, y por lo tanto el agua en los organismos es decididamente impuble.En consecuencia, si los organismos se colocan en agua pura, el agua se difunde en ellos. La afluencia de agua tiene dos resultados potenciales: uno es químico, los solutos celulares se diluyen tanto que se impide el funcionamiento normal; el segundo es mecánico, la afluencia de agua puede hacer que el organismo se hinche y finalmente estalle. Sin embargo, si el organismo está encerrado en un recipiente rígido, la afluencia de agua presuriza al organismo y el flujo de agua cesa con solo cambios modestos de volumen. Otra posibilidad que permite que los organismos vivan agua 'fresca' (es decir, pura) es tener mecanismos que permitan que el agua sea expulsada del organismo a la misma velocidad que ingresa.
Los problemas asociados con la difusión del agua hacia los organismos se eliminan si la pureza del agua es la misma afuera que dentro del organismo, y hay una serie de organismos (muchos animales marinos, algunos protistas) que requieren un ambiente tal donde la pureza del citosol es comparable a el del agua de mar.
También hay hábitats, denominados salinos ('salinos', aunque la sal no necesita ser NaCl) que tienen una pureza de agua incluso menor que la de la mayoría de los organismos. La mayoría de las formas de vida están excluidas de dichos ambientes salinos porque la difusión hace que pierdan agua y no pueden tolerar la disminución del contenido interno de agua que resulta de la pérdida de agua. Los organismos que son capaces de tolerar tales ambientes lo hacen al tener más de la concentración normal de solutos en su citosol, generalmente agregando solutos únicos que normalmente no se encuentran en los seres vivos.
Los problemas de vida en ambientes salinos no se limitan a reducir el contenido de agua. Los problemas adicionales provienen de las concentraciones tóxicas de los solutos que hacen que el hábitat sea salino. Generalmente, esto serían iones de sodio y cloruro pero hay hábitats donde otros solutos (por ejemplo, potasio, calcio) son dañinos. A excepción del sodio, que la mayoría de las plantas no requieren, estos elementos son esenciales para los seres vivos, pero a altas concentraciones, pueden interferir con el funcionamiento normal celular u organizativo y volverse tóxicos.
Oxígeno y Ambientes Acuáticos
El oxígeno es de importancia clave en los ambientes acuáticos. El oxígeno se disuelve fácilmente en el agua, pero su solubilidad es tal que el oxígeno generalmente está menos disponible que en los hábitats terrestres. De manera más significativa, su abundancia varía mucho más en los hábitats acuáticos que en los terrestres. Al considerar los ambientes terrestres, debido a la mezcla conveccional (vientos) debido a la menor densidad de aire en comparación con el agua, y, en menor medida, debido a la tasa mucho más rápida de difusión en los gases, los niveles de oxígeno del aire rara vez cambian mucho. Los procesos (principalmente los bióticos) pueden aumentar o disminuir los niveles de oxígeno, pero la mezcla rápida con el enorme reservorio de oxígeno en la atmósfera en su conjunto, que es ligeramente inferior al 20% de oxígeno, devuelve rápidamente el aire en estas áreas a los valores atmosféricos.
En contraste, el agua líquida es mucho más densa, su movimiento es mucho más lento y en consecuencia, es mucho más probable la posibilidad de áreas localizadas con concentraciones de oxígeno sustancialmente alteradas de las del hábitat en su conjunto. Bajo ciertas situaciones, los hábitats acuáticos estarán saturados de oxígeno, es decir, conteniendo todo el oxígeno que pueda disolverse en el agua, cantidad que depende de la temperatura y disminuye con el aumento de la temperatura. Como sería de esperar, el agua se satura de oxígeno cuando el aire y el agua están en contacto cercano (es decir, la superficie de cuerpos de agua) o en situaciones donde las tasas fotosintéticas son altas, aumentando así los niveles de oxígeno (generalmente estas áreas también están cerca de la superficie). Sin embargo, hay muchas situaciones acuáticas donde la cantidad de oxígeno disuelto no está en saturación. Generalmente, esto se debe al agotamiento de oxígeno por parte de los seres vivos y a la falta de mezcla con el agua que está en contacto con el aire, situación que se encuentra en el fondo de los lagos, en arroyos de movimiento lento, y suelos anegados. Los impedimentos para la mezcla harán que el agotamiento de oxígeno sea más probable. Dos de los impedimentos más comunes son los gradientes de densidad y los materiales sólidos. Los pantanos de Sphagnum suelen tener agua empobrecida en oxígeno a profundidades muy poco profundas, profundidades donde los niveles de oxígeno suelen ser mucho más altos. Esto es el resultado de que el tapete de esfagno dificulta el movimiento de la columna de agua. Aunque la atmósfera rica en oxígeno está a centímetros de distancia, los niveles de oxígeno bajan a casi cero. Algo similar puede suceder en estanques que están cubiertos con una gruesa 'basura de charco' de algas. Otro obstáculo significativo para el movimiento del agua son los gradientes de densidad establecidos por la temperatura. El agua menos densa 'flota' en agua más densa y esto reducirá la transferencia de oxígeno de la atmósfera al agua más densa. La densidad del agua disminuye a medida que la temperatura aumenta desde los 4 C; de ahí que en este rango, el agua más cálida sea menos densa que el agua más fría. Una consecuencia de esto es que el agua es estratificada ('estratificada') durante las épocas del año en que se calienta desde arriba, durante el verano en latitudes septentrionales, durante todo el año en los trópicos. El viento puede causar turbulencia en la columna de agua pero la penetración de esta turbulencia está limitada por las diferencias de densidad. Cuanto mayores son las diferencias de temperatura entre la parte superior e inferior, más resistente es el lago a la mezcla. Debido a esta estratificación, las capas inferiores de un lago pueden agotarse de oxígeno porque no se mezclan con el agua oxigenada en la superficie. En áreas tropicales, los bajos niveles de oxígeno en el fondo de los lagos ocurren durante todo el año, pero en áreas templadas, la mezcla y como resultado la aireación de las capas inferiores, se vuelve cada vez más probable durante la caída. Esto se debe a que, a diferencia del calentamiento del verano, que refuerza la estratificación al hacer que la parte superior sea más cálida, el enfriamiento descompone la estratificación al hacer que la superficie sea más densa. Mientras el lago esté en un lugar que se enfríe lo suficiente, el lago eventualmente se convertirá en 'isotérmico', es decir, la misma temperatura en toda la columna de agua. Al ser isotérmico también hace que el lago tenga la misma densidad de arriba a abajo y, siempre y cuando el viento sople lo suficientemente fuerte (qué tan fuerte depende de la profundidad del lago), el lago se 'mezclará', trayendo agua aireada de abajo hacia arriba. Esto es significativo desde el punto de vista nutricional porque el material orgánico se asienta en el fondo del lago y se descompone allí. Los minerales liberados por descomposición solo se distribuyen por toda la columna de agua cuando se mezcla el lago. El enfriamiento adicional del lago vuelve a resultar en estratificación, ya que la densidad del agua disminuye (y el agua más fría flota en la parte superior) a medida que la temperatura desciende de 4 a 0 C. Si el lago se congela, la adición de una capa de hielo elimina cualquier mezcla impulsada por el viento y los niveles de oxígeno en el fondo del lago una vez más caer si la respiración excede la fotosíntesis.Esto suele ser el caso (aunque alguna fotosíntesis sí se realiza en lagos cubiertos de hielo, es relativamente baja debido a la disminución de la penetración de la luz). En la primavera (en el hemisferio norte) el calentamiento de la superficie primero elimina el hielo y luego elimina la estratificación, haciendo que el lago sea uniforme en temperatura y densidad a 4 C, y permitiendo nuevamente la mezcla por el viento. El calentamiento adicional hace que el lago vuelva a ser estratificado. Así, los lagos en hábitats templados tienen un ciclo anual que incluye dos breves periodos en otoño y primavera donde es muy probable que se mezclen, siempre y cuando el viento sople y el lago no sea demasiado profundo. Esta situación, denominada “rotación” de primavera y otoño, es muy significativa porque permite mezclar la columna de agua, llevar agua oxigenada al fondo del lago (y también llevar minerales a la parte superior del lago (ver abajo)).
Ambientes ligeros y acuáticos
Como es el caso en los ambientes terrestres, la luz juega un papel importante en dictar la distribución de los organismos fotosintéticos y es aún más importante en los ambientes acuáticos porque pueden desarrollarse diferencias significativas en las concentraciones de oxígeno dentro de la columna de agua, dependiendo del equilibrio entre fotosíntesis y respiración. La intensidad de la luz está influenciada por la profundidad y transparencia del agua, esta última a menudo está fuertemente influenciada por el número de seres vivos que viven en el agua. Dado que los fotoautótrofos acuáticos necesitan luz para sobrevivir, necesitan estar en los niveles superiores del agua, o, en el caso de algunas plantas con flores, flotar en la superficie o incluso emerger de ella. El fitoplancton (pequeños organismos fotosintéticos que 'flotan' en la columna de agua) necesitan tener características que impidan su sedimentación. Las características que a veces son significativas son grandes proporciones de superficie a volumen (es decir, no esferas), cuerpos de petróleo o aire que disminuyen la densidad, flagelos y fototaxis (movimiento hacia la luz (ver criptófitas). Se sabe que algunos fitoplancton migran hacia arriba y hacia abajo diurnalmente, moviéndose hacia arriba para ganar más luz durante las horas del día.
Corrientes y Ambientes Acuáticos
Las corrientes son muy significativas en algunos ambientes acuáticos y pueden tener una fuerte influencia en la estructura de la comunidad. Las corrientes son significativas porque pueden mezclar porciones de la columna de agua, en particular, pueden llevar oxígeno de las capas superiores a las capas inferiores y nutrientes de las capas inferiores a las capas superiores. Esto es significativo porque el oxígeno puede ser limitante en las capas inferiores y los nutrientes pueden ser limitantes en las capas superiores. En consecuencia, la corriente puede cambiar sustancialmente las actividades que se llevan a cabo.
Nutrientes y Ambientes Acuáticos
Los nutrientes suelen limitar la productividad primaria y, en consecuencia, la actividad total en situaciones acuáticas. Esto es resultado de que la mayoría de los ambientes acuáticos tienen dos partes distintas: las capas superiores donde se produce la fotosíntesis y donde se incorporan nutrientes inorgánicos a la biomasa, y la región inferior, donde normalmente hay poca fotosíntesis, pero donde la biomasa de las capas superiores tiende a se asientan, se produce la descomposición y se liberan nutrientes. En tal sistema las actividades tanto de la parte superior como de la inferior pueden verse limitadas por la falta de interacción entre las dos partes: las capas superiores están limitadas por la falta de nutrientes, de ahí que se produzca poca acumulación de biomoléculas; la capa inferior se ve limitada por la falta de material para comer.
Los nutrientes transportados a los sistemas acuáticos pueden ser muy importantes. Los nutrientes pueden ser transportados a los sistemas acuáticos por escorrentía de la tierra circundante, especialmente tierras agrícolas donde se agregan nutrientes a través de la fertilización. El nitrógeno y el fósforo a menudo juegan un papel clave en la determinación de la cantidad de producción primaria que ocurre y la cantidad total de actividad biológica que tiene lugar. Sin embargo, como fue el caso en los sistemas terrestres, existen circunstancias en las que otros nutrientes pueden desempeñar un papel clave.