
1.30: Amenazas a la agricultura- insectos y patógenos
- Page ID
- 56458

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)

Como cualquier agricultor te dirá hay muchas amenazas para el cultivo de plantas. Algunos de estos han sido cubiertos en capítulos anteriores: extremos en temperatura y humedad (Capítulo 26); e interacciones con otras plantas, es decir, malezas (Capítulo 29). Un gran número de amenazas adicionales provienen de interacciones tróficas con una vasta diversidad de pastores, parásitos y depredadores. Ciertamente los más familiares de estos son los insectos pero otros invertebrados (ácaros, nematodos, moluscos) y vertebrados pueden ser obstáculos importantes para el crecimiento de las plantas. Además, una gran cantidad de hongos, bacterias (Figura 1), protistas y virus también son amenazas importantes.
TEMAS
- Herbívoros en pastoreo
- Herbívoros absorbentes
- Defensas vegetales
- Límite
- Productos Químicos
- Fenología
- Una muestra de enfermedades de las plantas
- Fitoplasmas y enfermedad definitoria
- Tizón tardío de la papa: el triángulo de la enfermedad
- Enfermedad de la roya del trigo: ciclos de vida de patógenos complejos
- Enfermedad del tizón de maíz: trufas mexicanas
- Enfermedad del olmo holandés: la reacción exagerada puede matarte
- Enfermedades por explosión del arroz: estrategias que cambian
- Tiña del fuego de fitopatógenos bacterianos de manzana
- Enfermedad de la hiel de la corona: hacer de una enfermedad una herramienta
- Enfermedades víricas: virus mosiaco del tabaco, virus del mosaico de la coliflor, enfermedad de la mancha anular de la papaya
Herbívoros en pastoreo
La mayoría de los herbívoros animales, por ejemplo, las marmotas y los escarabajos japoneses adultos, son considerados pastoreros, comiendo partes de la planta y utilizando su movilidad para llegar a otras plantas. Sin embargo, un número sustancial de insectos herbívoros tienen etapas de alimentación (generalmente larvas, por ejemplo, min e rs de hoja, barrenadores de ceniza esmeralda, escarabajos larvarios de la papa) que son inmóviles hasta llegar a nuevas plantas hospedadoras. Estos podrían ser mejor considerados parásitos porque se alimentan únicamente de una planta hospedadora. La mayoría de los herbívoros no animales (hongos, bacterias) son básicamente inmóviles y se consideran parásitos aunque algunos son capaces de crecer de un huésped a otro y muchos son transportados de una planta a otra por el viento, la lluvia y los insectos.

La mayoría de los pastoreros de las plantas son ingentes (ver Capítulo 27), retiran y consumen parte de la planta, la trituran hasta cierto punto y después digieren las piezas en un tubo interno, y finalmente excretan indigeribles a su entorno. Los pastoreros, aunque pueden reducir severamente los rendimientos de los cultivos, rara vez matan por completo a la planta que está comiendo tanto porque tienden a especializarse solo en parte de la planta (más comúnmente hojas) como porque la planta es capaz de reemplazar los órganos/partes de órganos que se han perdido.
El material vegetal adquirido por los pastoreros generalmente no es muy denso en nutrientes, pero varía sustancialmente dependiendo de la parte de la planta consumida. La calidad nutricional está influenciada en gran medida por la cantidad de células vivas presentes (las células muertas son básicamente todas las paredes celulares y son indigeribles para la mayoría de los herbívoros). La mayoría de los herbívoros se especializan en uno de los tejidos enumerados a continuación, pero hay pocos generalistas que se alimentan de múltiples tejidos, por ejemplo, los navegadores como los alces y los ciervos a menudo comen tallos, hojas y meristemos.
- crecimiento primario de tallos y raíces — aunque los tallos y raíces en crecimiento primario tienen más células vivas que las que tienen crecimiento secundario, siguen siendo pobres en nutrientes. La mayoría de esas células que están vivas no son muy metabólicamente activas y, en consecuencia, son más pobres en nutrientes en comparación con las células que son más metabólicamente activas (por ejemplo, células del mesófilo foliar).
- tallos y raíces leñosas, duramen: El duramen no tiene células vivas y tiene mayores niveles de químicos secundarios. Generalmente tiene la menor calidad nutricional de cualquier tejido vegetal, pero hay especialistas en duramen
- tallos y raíces leñosas, madera de savia: La albura tiene algunas células vivas (los rayos) y estas pueden estar 'cargadas' con almidón y otros nutrientes, dependiendo de la temporada. De ahí que la albura sea considerablemente más nutritiva que el duramen, pero todavía está compuesta en gran parte de material de la pared celular.
- tallos y raíces leñosas, cambium vascular: este tejido meristemático es rico en nutrientes y tiene relativamente menos material de la pared celular en comparación con las células maduras. Además, es más extenso que los meristemos apicales, es decir, hay más material disponible para comer.
- el tejido de corteza de tallos leñosos es pobre en nutrientes con pocas células vivas y abundancia de químicos secundarios. La porción de células que están vivas disminuye a medida que el tallo/raíz envejece y por lo tanto la corteza de los tallos más jóvenes es más nutritiva que la corteza de los tallos más viejos
- tallos y raíces que han sido modificados como estructuras de almacenamiento — rizomas, cormos, bulbos, estolones. Están compuestas por células vivas que están almacenando carbohidratos, generalmente en forma de almidón. Son ricos en carbohidratos digeribles pero pueden ser deficientes en aminoácidos (proteínas) y otros nutrientes
- Alimentadores de floema: Estos organismos (incluyendo pulgones, tolvas foliares, chinches harinosos y moscas blancas, todas plagas importantes de la agricultura) no están consumiendo ninguna célula, sino que aprovechan el sistema de transporte del floema y adquieren los materiales que se transportan a través de él. La savia del floema tiene concentraciones muy altas de sacarosa junto con concentraciones mucho más pequeñas de aminoácidos y algunos minerales. Debido a que la proporción de carbohidratos a aminoácidos en la savia del floema es tan alta que la mayoría de los alimentadores de floema excretan 'melaza', esencialmente savia de floema, todavía rica en carbohidratos pero con la mayoría de los aminoácidos eliminados. La melaza es utilizada por varios otros organismos, en particular algunas especies de hormigas y el hongo del moho hollíneo.
- Alimentadores de xilema: La savia de xilema tiene muy pocos nutrientes, casi sin carbohidratos pero con algunos iones minerales. Además, mientras que los alimentadores de floema adquieren savia pasivamente porque está bajo presión dentro de las células del floema, la savia de xilema está bajo tensión y requiere 'succión' (desarrollando mayores tensiones que las ya presentes) para que fluya hacia los alimentadores de xilema. Sin embargo, hay algunos insectos (cigarras larvarias, chinches larvarias) que se alimentan de xilema.

Herbívoros absorbentes
A diferencia de los pastoreros, la mayoría de los herbívoros vegetales no animales son absorbentes (ver Capítulo 27), absorbiendo nutrientes de 'su entorno', siendo 'su entorno' el interior de una planta. La mayoría de estos organismos son bacterias y hongos, pero algunos, como los mohos de agua, son protistas y algunos son animales (por ejemplo, nematodos, las etapas larvarias de algunos insectos). La mayoría de estos organismos se consideran parásitos y generalmente se consideran patógenos de plantas. Invaden los tejidos vegetales y luego adquieren materiales de una de tres maneras: (1) el parásito se alimenta de materiales que 'se filtran' de las células hospedadoras. (2) el parásito mata las células hospedadoras, haciendo que los materiales celulares estén disponibles. Los organismos que alimentan esto se describen como necrotróficos (alimentándose de material muerto). (3) el parásito y el hospedador forman una estructura llamada haustorio, una fusión tanto de la membrana de la célula huésped como de la membrana celular del parásito. (Obsérvese que el desarrollo del haustorio generalmente requiere la ruptura de las paredes celulares tanto del huésped como del parásito). Los materiales se transfieren de hospedador a parásito a través del haustorio y tanto el huésped como el parásito permanecen vivos. Los organismos que se alimentan de esta manera se describen como biotróficos (es decir, alimentándose de material vivo). Algunos biótrofos son herbívoros relativamente benignos (se benefician de mantener vivo a su huésped) y, en consecuencia, muchos adquieren cantidades relativamente pequeñas de recursos de sus huéspedes. Pero muchos biótrofos (por ejemplo, oxidas, tizones, mildiú velloso, mildiú polvoriento) pueden ser devastadores para los agricultores ya que las infestaciones a menudo pueden reducir los rendimientos de las plantas a casi nada.

Defensas vegetales
Un límite efectivo:
A excepción de los grandes animales de pastoreo, los organismos necesitan meterse dentro de los tejidos de las plantas para alimentarse de la planta. El límite lo hace difícil al producir una superficie difícil de penetrar e inhóspita para la vida (ver Capítulo 3). Para el crecimiento primario por encima del suelo, el límite consiste en células epidérmicas que están estrechamente unidas entre sí, dificultando la penetración entre las células. Adicionalmente, el crecimiento primario sobre el suelo está recubierto con una cutícula que es difícil de perforar y difícil de vivir porque es hidrofóbica. Bajo tierra, las raíces primarias maduras tienen una endodermis que cumple muchas de las mismas funciones que la epidermis + cutícula hace sobre el suelo. Sin embargo, las partes más jóvenes de las raíces no han desarrollado una endodermis. Esto facilita la adquisición de agua pero también facilita la entrada de patógenos.
Los patógenos/herbívoros ganan entrada tanto al utilizar aberturas estomáticas como también al tener partes bucales que pueden penetrar la cutícula/epidermis, o tener un tipo de aguja o sierra, como ovipositer (órgano ponedor de huevos) o, en el caso de algunos hongos, produciendo un appresorio, un tipo celular especializado que es capaz de fusionarse con la epidermis y producir una estructura que pueda penetrar en ella. Los patógenos y herbívoros también ingresan a través de heridas y aberturas en la epidermis como resultado de procesos de crecimiento, por ejemplo, raíces de ramas, desprendimiento de hojas y ramas.
Química:
Todas las plantas producen diversos químicos que influyen en herbívoros y patógenos. Algunos están alimentando químicos disuasorios que son venenosos o pueden anunciar que los venenos están presentes dentro de la planta. Al mismo tiempo, algunos químicos producidos por las plantas atraen claramente a ciertos herbívoros. El supuesto es que estos químicos alguna vez habían servido como disuasores de alimentación pero que el herbívoro ha desarrollado medios para desintoxicar el veneno y ahora está utilizando el químico para identificar un huésped que relativamente pocos competidores podrán utilizar. Por ejemplo, la mayoría de los insectos herbívoros no comerán algodoncillo pero varios insectos (larvas de mariposa monarca, el escarabajo del algodoncillo y los insectos algodoncillo) eligen alimentarse de algodoncillo. Algunos incluso utilizan los venenos de la planta como propios, haciéndolos menos propensos a ser consumidos.
Las plantas también producen productos químicos porque reconocen que están bajo ataque. La producción de estos químicos 'inducidos' implica que la planta tiene la capacidad de detectar la presencia del patógeno y los químicos producidos pueden: (1) matar al invasor (fitoalexinas), (2) desencadenar respuestas defensivas en plantas vecinas y/o atraer depredadores que puedan controlar al herbívoro, o (3) provocar una “respuesta hipersensible”, provocando que el tejido invadido muera rápidamente. Tenga en cuenta que la respuesta hipersensible puede ser efectiva para disuadir a los patógenos biotróficos pero en realidad beneficia a los patógenos necrotróficos.
Fenología:
Las plantas (y los agricultores) pueden evitar los herbívoros/depredadores ajustando el momento de germinación de las semillas, crecimiento y producción de flores/frutos.
A Muestreo de enfermedades de las plantas:
Poinsettias, fitoplamas y la naturaleza de las enfermedades de las plantas
La flor de pascua no siempre se veía como la mayoría de ellas ahora. La planta en estado silvestre y la planta que originalmente se propagó como planta ornamental era planta mucho más alta y producía relativamente pocas 'flores' (las estructuras que parecen una flor son en realidad un racimo de flores rodeadas de brácteas coloridas, generalmente rojas). A principios del siglo XX apareció una planta con una forma novedosa, una que era mucho más corta, se ramificó con mucha más frecuencia y producía más flores. Esta forma podría propagarse por esquejes y el supuesto era que la planta original era un mutante, un 'deporte', como la naranja naval original. El deporte generalmente no puede ser perpetuado por semilla porque las mutaciones que los hacen distintivos suelen ser recesivas. Pero a menudo pueden perpetuarse por esquejes porque los esquejes son esencialmente crecimiento continuo de la planta original. Hay otros fenómenos además de las mutaciones que podrían explicar un patrón de crecimiento alterado que puede perpetuarse por esquejes. Dos posibilidades son los virus y los fitoplasmas (pequeñas bacterias que carecen de paredes celulares que viven solo en células vegetales). Tenga en cuenta que estas tres posibilidades para la planta anormal (mutación, infección viral infección por fitoplasma) todas podrían describirse como “enfermedad de la planta” pero que lo inusual es que la planta enferma es deseable no indeseable. ¡Resulta que casi toda la flor de pascua que se produce hoy están enfermas! Y el culpable resulta ser un fitoplasma.

Entonces, ¿qué es la enfermedad vegetal? En la discusión de nochebuena se asoció con 'plantas anormales'. Una dificultad con esta definición es que es 'normal' que las entidades biológicas sean 'anormales'. Es decir, si nos fijamos en una población (grupo de organismos) no todos son iguales, varían, algunos están 'fuera de la norma'. Adicionalmente, 'normal' no se define fácilmente. Se puede definir estadísticamente como una tendencia central, por ejemplo anormal es una desviación estándar más de una media. Aunque no es perfecta, la definición 'anormal' funciona con plantas en parte porque las plantas bajo consideración son típicamente las de importancia económica y 'normal' es un tipo de planta que proporciona el mayor rendimiento económico. Adicionalmente, la mayoría de los cultivos han sido criados para ser uniformes.
Así, una planta enferma es reconocida por una estructura o funcionamiento anormal. Los síntomas comunes de las plantas enfermas incluyeron crecimiento atrofiado o deformado y hojas amarillentas (cloróticas) o muertas (necróticas). Pero ocasionalmente se puede encontrar una planta enferma que es 'más grande de lo normal', que se ramifica más de lo normal, o que tiene hojas que persisten en la planta longe r de lo normal. De manera arbitraria, la enfermedad vegetal excluye a los insectos herbívoros aunque tengan similitudes con los patógenos de plantas “verdaderos”. Dado que este libro trata sobre la vida inanimada no consideraremos a los insectos herbívoros excepto como se introdujo anteriormente y como se cubre en el siguiente capítulo que trata sobre el desarrollo de nuevas plantas.
Tizón tardío de la papa y el 'triángulo de la enfermedad '

Las causas de funcionamiento anormal, es decir, plantas enfermas, son legiones, incluyendo: condiciones climáticas, factores nutricionales (suelo), cambios genéticos y un amplio espectro de trastornos resultantes de interacciones con una variedad de otros organismos: bacterias, hongos, mohos de agua, nematodos, insectos y con biótico entidades como virus. Con respecto a estas causas bióticas de enfermedades vegetales, los fitopatólogos describen lo que se llama el 'triángulo de enfermedad' donde la enfermedad es el resultado de una combinación de condiciones ambientales, susceptibilidad del huésped y virulencia del organismo causante de enfermedades. La enfermedad es consecuencia de que una planta susceptible se encuentra con un patógeno competente (es decir, virulento) bajo condiciones ambientales que favorecen la invasión y propagación del patógeno. Un ejemplo clásico del triángulo de enfermedades es el tizón tardío de la papa, causado por el moho de agua Phytophthora. La papa es originaria de América del Sur pero fue traída a Europa en el siglo XVII como cultivo alimentario. El organismo de la enfermedad, que también afecta a los tomates, aparentemente se originó en México a principios del siglo XIX. La enfermedad requiere condiciones frescas y húmedas, un huésped susceptible de papa y un patógeno virulento. El patógeno se extendió por toda América del Norte a principios del siglo XIX y llegó a Europa probablemente como resultado de la importación de papas enfermas a Bélgica en 1845. Rápidamente se extendió por toda Europa, y en particular a Irlanda, donde las papas eran la principal fuente de alimento y se cultivaban extensamente en monocultivos. Las condiciones frías y húmedas típicas de Irlanda fueron adecuadas para el crecimiento y reproducción del patógeno. Además, las papas cultivadas fueron genéticamente uniformes y también altamente susceptibles al tizón. El resultado fue devastador, más de un millón de irlandeses murieron de inanición y otro millón emigró, en su mayoría a Estados Unidos. La severidad del tizón varió en los próximos veinte años, principalmente debido a las fluctuaciones en las condiciones ambientales. Tenga en cuenta que el fracaso de los cultivos no solo elimina los alimentos, sino que también elimina las 'patatas de semilla' (papas guardadas para ser plantadas al año siguiente). Así, aunque las condiciones favorables pudieran producir un cultivo relativamente alto (rendimiento por acre), la escasez de alimentos persiste porque es posible que se hayan plantado menos acres. El tizón tardío de la papa sigue siendo un problema, con brotes ligados a las condiciones ambientales. Existen variedades parcialmente resistentes pero éstas son solo temporales porque el patógeno evoluciona para superar la resistencia de la planta.

El patógeno es un parásito biotrófico obligado, lo que significa que solo puede sobrevivir en un huésped vivo. Sobrevive año en año en tubérculos de papa o plantas de papa que quedan en el campo. Se propaga fácilmente por esporas (generalmente asexuales) que requieren humedad para mantenerse con vida y son dispersadas por el viento y por las gotas de lluvia. El crecimiento y la propagación de patógenos pueden ser extremadamente rápidos si las condiciones climáticas son apropiadas. Las esporas germinan rápidamente y entran en las plantas susceptibles a través de estomas, heridas y directamente a través de la cutícula. Una vez dentro de la hoja producen hifas que crecen entre las células y producen haustoria que penetran en la pared celular e interactúan con la membrana celular hospedadora y permiten que los nutrientes pasen al hongo.
La enfermedad requiere una coincidencia entre el huésped y el patógeno. P. infestans infecta papa y también tomate, que es un pariente cercano de la papa. Pero muchos otros parientes cercanos no son anfitriones adecuados. Y hay otras especies de Phytophthora que afectan a otras especies de plantas pero no infectan a las papas, por ejemplo P. quercina provoca el síndrome de muerte súbita del roble
Roya del trigo : ciclos de vida complejos del parásito
Hay muchas enfermedades diferentes de la roya que afectan a una variedad de huéspedes, algunos no utilizan dos hospedadores diferentes, pero todas son específicas del huésped, lo que significa que solo pueden afectar a un grupo de plantas estrechamente relacionadas: una variedad, una especie o varias especies de un solo género. El ciclo de vida de la roya del trigo se introdujo en el Capítulo 12 y a continuación se reproduce una cifra de ese capítulo. Tenga en cuenta que se producen cinco tipos diferentes de esporas que difieren entre sí por su nivel de ploidía (diploide, dikaryon o haploide), en qué tipo de planta se producen, y en qué tipo de planta pueden crecer (si la hay) y cómo funcionan.

Las teliosporas son esporas de dikaryon y la única etapa del ciclo de vida que crece, aunque de manera muy temporal, independientemente de un hospedador. También es la única etapa que puede pasar el invierno.
Las basidiosporas son esporas haploides producidas después de germinar la teliosporas y someterse a cariogamia para producir una célula diploide que luego se somete a meiosis para producir basidiosporas haploides. Estas esporas se dispersan en el aire y solo germinan y crecen en plantas de agracejo.
Las pinciosporas son esporas haploides producidas a partir de hifas producidas en una estructura que emerge de la superficie superior de las hojas de agracejo. También en esta estructura se encuentran hifas receptivas haploides que pueden recibir pinciosporas, fusionarse con ellas (plasmogamia) para formar hifas dikaryon. Estas crecen hasta la superficie inferior de la hoja y forman una estructura llamada aecium que produce aeciosporas de dikaryon que son dispersadas por el viento e infectan las plantas de trigo.
El crecimiento micelial dicótico de las aeciosporas puede causar daños significativos a la planta hospedadora de trigo y también produce urideosporas, producidas en estructuras anaranjadas llamadas uremia. Las uridosporas propagaron Puccinia graminis a otras plantas de trigo, facilitando la propagación de enfermedades a través de los cultivos de trigo.
A medida que la planta de trigo comienza a sentir, tanto por el patógeno como a partir de su ciclo natural, monocárpico, las hifas dicariotas en la planta de trigo producen teliosporas, esporas que pueden sobrevivir al invierno y completar el ciclo de vida.

Debido a que el trigo es un cultivo extremadamente importante y debido a que la enfermedad se propaga rápidamente y evoluciona rápidamente, la roya del trigo es probablemente la enfermedad agrícola más significativa. La resistencia a la enfermedad involucra genética tanto del hongo como de la planta. Las plantas resistentes tienen la capacidad de reconocer la invasión por el hongo y responder a ella. El reconocimiento del hongo es el resultado de que la planta percibe un químico producido por el hongo está presente. Un hongo avirulento puede volverse virulento al volverse irreconocible, típicamente al no producir una sustancia química específica que los hongos avirulentos (y por lo tanto reconocibles) habían estado fabricando. De ahí que la virulencia en el patógeno sea típicamente recesiva (incapacidad para hacer algo que el tipo salvaje haga). La resistencia de las plantas requiere un gen dominante que le dé a la planta la capacidad de reconocer y responder a la presencia del virus.
Smut de maíz — Trufas mexicanas

El tizón de maíz produce agallas grandes y distintivas que son comestibles y altamente deseables en México. En otras partes del mundo la enfermedad es muy poco bienvenida, reduciendo sustancialmente los rendimientos de los cultivos. Al igual que las oxidas, las enfermedades del tizón son basidiomicetos y al igual que las oxidas el hongo generalmente se encuentra en el estado dicariónico, con células que poseen dos núcleos haploides. Los basidiomicetos solo producen una sola célula diploide que inmediatamente se somete a meiosis para formar esporas haploides. Estas germinan y crecen hasta convertirse en hifas haploides que en los tizones pueden producir nuevas células por gemación (como la levadura) y como la levadura se puede cultivar en un medio artificial, lo que significa que esta etapa NO es biotrófica sino que actúa como un saprófito. Sin embargo, cuando las hifas haploides de dos tipos de apareamiento diferentes se encuentran entre sí y se fusionan y forman una hifa dicariota, ahora es biotrófica, solo capaz de sobrevivir en células vivas de plantas especificas, en este caso, maíz y teosinte (un pariente cercano del maíz). De ahí que para ser un patógeno exitoso las hifas haploides necesitan encontrarse en material de maíz vivo (o teosinte). Adicionalmente, las hifas dikaryon solo invaden los tejidos activos, más comúnmente las flores, donde inducen crecimiento anormal, agallas, con células muy agrandadas rodeadas de hifas. No forman haustoria típica sino que forman estructuras que permiten la transferencia de materiales de la planta al hongo. A medida que las agallas maduran cambian de color de blanco sedoso a negro y también cambian las texturas, volviéndose más suaves. Aunque las agallas se ven 'fúngicas' (¡si tal cosa es posible!) en realidad son en su mayoría tejido vegetal. Dentro de las agallas algunas células fúngicas se someten a cariogamia (fusión de los dos núcleos haploides) para formar una célula diploide y completar el ciclo sexual. El tizón de maíz es la enfermedad de carbón más importante, pero hay otros hongos de tizón que infectan otras hierbas agrícolas (caña de azúcar, cebada, avena) así como los tizones que atacan pastos y juncias silvestres.
Enfermedad por explosión del arroz : estrategias de patógenos cambiantes

La explosión del arroz es una enfermedad fúngica devastadora que afecta a varios cereales pero es más significativa en afectar al arroz. Es un ascomiceto y la etapa dikaryon es breve en tiempo y extensión. El patógeno se describe técnicamente como un hemibiótrofo porque inicialmente se comporta como un biótrofo, sobreviviendo con células vegetales vivas, pero eventualmente se convierte en necrótrofo, matando células y obteniendo nutrientes de ellas. Las esporas haploides aterrizan sobre hojas de arroz, frutos en desarrollo y otras partes de la planta y son capaces de penetrar en la cutícula generando presiones sustanciales hidrostáticamente dentro de una célula especializada llamada appressorium. Una vez dentro del hongo es capaz de propagarse ingresando células vegetales individuales a través de plasmodesmas. Después de una cierta cantidad de tiempo que depende del tejido que ha sido invadido, el hongo cambia a un estilo de vida necrotrófico, matando a las células hospedadoras.
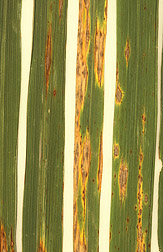
Enfermedad del olmo holandés : exagerar puede ser mortal
La enfermedad del olmo holandés es causada por un hongo ascomiceto que tiene una relación simbiótica con un escarabajo de la corteza, un tipo de escarabajo que se alimenta del cámbium vascular y los tejidos (xilema secundario y floema) que produce el cambium vascular. El hongo es transportado de árboles infectados a árboles nuevos por el escarabajo y se beneficia no solo por el transporte sino por ser colocado dentro de la planta en los tejidos de los que se alimenta el hongo. El hongo se alimenta únicamente de células moribundos. pero, desafortunadamente para el árbol, su presencia desencadena la producción de gomas. Presumiblemente tal respuesta podría ayudar a detener la propagación del hongo al dificultar su movimiento. Sin embargo, en los olmos las sustancias que se producen taponan los vasos y traquieds del hospedero, haciéndolos incapaces de transportar agua hasta la planta. Al igual que una serie de enfermedades humanas, el daño de un patógeno proviene principalmente de la respuesta del huésped y menos de la actividad real del patógeno. La enfermedad del olmo holandés es una de las muchas “enfermedades de marchitez” causadas tanto por hongos como por bacterias que resultan en una reducción del transporte de agua y marchitamiento. En la mayoría de estos el bloqueo del tejido del xilema es el resultado de la respuesta de la planta al patógeno.

Tizón de fuego y fitopatógenos bacterianos
La mayoría de los patógenos de las plantas son fúngicos, pero algunos, como el tizón tardío de la papa, son oomicetos y algunos, como los fitoplasmas de las poinsettias son bacterias. Si bien los fitoplasmas son inusuales porque son parásitos intracelulares obligados, la mayoría de las enfermedades bacterianas no ingresan a la célula sino que producen moléculas 'efectoras' que son capaces de transferir a las células vivas y efectuar resultados específicos como respuestas hormonales que provocan tumores, exudación de materiales, o muerte celular. La mayoría de los patógenos bacterianos de plantas no se clasifican fácilmente como biótrofo/necrótrofo aunque claramente los fitoplasmas son biótrofos, al igual que el Agrobacterium formador de agallas (ver abajo). El tizón del fuego es interesante a este respecto. Es causada por Erwinia amylovora que afecta a miembros de la familia de las rosas, en particular manzanas y peras, a veces con resultados catastróficos. La bacteria puede vivir como una epífita inofensiva que vive de azúcares exudados por los estigmas de las flores y los nectarios en la base de las flores. De estas ubicaciones E. amylovora puede ser propagada por polinizadores a lo largo de una planta y a lo largo de todo un huerto. En algún momento se activa para convertirse en un asociado mucho menos benigno y se vuelve necrógena, matando a las células hospedadoras como consecuencia de las moléculas efectoras que produce.

Enfermedad de la vesícula de la corona : una enfermedad utilizada como herramienta contra la enfermedad

Muchas enfermedades de las plantas alteran el patrón de crecimiento de las plantas, a menudo produciendo estructuras características llamadas agallas. El crecimiento anormal que se manifiesta en una agalla suele ser consecuencia de cambios en las cantidades de reguladores del crecimiento de las plantas, también conocidas como hormonas. Esto puede ser el resultado del patógeno que produce estos químicos (de hecho, una hormona vegetal, el ácido giberélico, se descubrió por primera vez como resultado de su producción por una enfermedad fúngica). Pero los niveles alterados de sustancias de crecimiento vegetal también pueden ser consecuencia de que el patógeno haga que su huésped produzca más o menos de una sustancia en particular. La forma más común en que esto sucede es que el patógeno altera la expresión génica en su huésped. Agrobacterium tumifaciens hace esto alterando realmente los genes presentes en las células hospedadoras a través de un proceso llamado transformación, un tipo de transferencia génica horizontal. Entre otros genes que se transfieren se encuentra uno que codifica para la producción de la sustancia de crecimiento vegetal citoquinina que juega un papel en la proliferación celular requerida para la formación de agallas. La bacteria vive saprofíticamente en el suelo pero es capaz de reconocer plantas heridas cuando entra en contacto con ellas y se transforma en una forma virulenta que se abre paso en la planta. A. tumifacien s es un patógeno significativo en varias especies de cultivos, en particular las perennes como el nogal, el albaricoque y la ciruela. A. tumifaciens destaca por su muy amplia gama de hospedadores que es una de las razones por las que ha demostrado ser útil en la ingeniería genética de plantas ya que puede ser utilizada para transferir genes específicos de un organismo a otro (Capítulo 31).
Virus del mosaico del tabaco, virus del mosaico de la coliflor y enfermedad de la mancha anular de la papaya

Las plantas se ven afectadas por una gran cantidad de virus que pueden causar pérdidas de cosechas muy significativas. El virus del mosaico del tabaco (TMV) no es uno de los patógenos virales más dañinos pero es notable por su papel en la comprensión de los virus y su uso en la bioingeniería. El TMV fue el primer virus que se aisló y purificó y esto permitió el reconocimiento de que los agentes infecciosos no necesitan ser celulares para que causen enfermedades. Los virus están compuestos tanto de proteínas como de material genético (ya sea ADN o ARN) y al entrar en una célula insertan material genético que codifica para la producción de proteína viral y material genético. Una parte crítica del genoma viral insertado es una región llamada promotor que 'promueve' la expresión de los genes aguas abajo de él, asegurando así su expresión. Los ingenieros genéticos han utilizado esta región del genoma de varios virus con el fin de insertar genes deseables de un organismo en otro organismo. En particular, el virus del mosaico de la coliflor (CaMV) ha demostrado ser particularmente útil en la producción de organismos “modificados genéticamente”. Un cultivo donde el promotor CaMV fue muy útil fue en el desarrollo de cepas de papaya que son resistentes a la enfermedad de la mancha anular de la papaya, una enfermedad que es causada por otro virus más que amenazó con eliminar la papaya como cultivo comercial.