
3.1: Flujo de Energía a través de Ecosistemas
- Page ID
- 52802

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
Un ecosistema es una comunidad de organismos y su ambiente abiótico (no vivo). Los ecosistemas pueden ser pequeños, como las charcas de marea que se encuentran cerca de las costas rocosas de muchos océanos, o grandes, como las que se encuentran en la selva tropical de la Amazonía en Brasil (Figura\(\PageIndex{1}\)).
Existen tres amplias categorías de ecosistemas basados en su entorno general: agua dulce, marino y terrestre. Dentro de estas tres categorías se encuentran los tipos de ecosistemas individuales basados en el hábitat ambiental y los organismos presentes.
Los ecosistemas de agua dulce son los menos comunes, ocurriendo solo en 1.8 por ciento de la superficie de la Tierra. Estos sistemas comprenden lagos, ríos, arroyos y manantiales; son muy diversos y soportan una variedad de animales, plantas, hongos, protistas y procariotas.
Los ecosistemas marinos son los más comunes, comprenden el 75 por ciento de la superficie de la Tierra y constan de tres tipos básicos: océano poco profundo, agua oceánica profunda y fondo oceánico profundo. Los ecosistemas oceánicos poco profundos incluyen ecosistemas de arrecifes de coral extremadamente biodiversos. Pequeños organismos fotosintéticos suspendidos en las aguas oceánicas, conocidos colectivamente como fitoplancton, realizan el 40 por ciento de toda la fotosíntesis en la Tierra. Los ecosistemas de fondo marino profundo contienen una amplia variedad de organismos marinos. Estos ecosistemas son tan profundos que la luz es incapaz de alcanzarlos.
Los ecosistemas terrestres, también conocidos por su diversidad, se agrupan en grandes categorías llamadas biomas. Un bioma es una comunidad de organismos a gran escala, definida principalmente en tierra por los tipos de plantas dominantes que existen en regiones geográficas del planeta con condiciones climáticas similares. Ejemplos de biomas incluyen selvas tropicales, sabanas, desiertos, pastizales, bosques templados y tundras. Agrupar estos ecosistemas en solo unas pocas categorías de biomas oscurece la gran diversidad de los ecosistemas individuales dentro de ellos. Por ejemplo, los cactus saguaro (Carnegiea gigantean) y otra vida vegetal en el desierto sonorense, en Estados Unidos, son relativamente diversos comparados con el desolado desierto rocoso de Boa Vista, una isla frente a la costa de África Occidental (Figura\(\PageIndex{2}\)).
Cadenas Alimentarias y Redes Alimentarias

Una cadena alimentaria es una secuencia lineal de organismos a través de los cuales los nutrientes y la energía pasan a medida que un organismo se come a otro. Los niveles en la cadena alimentaria son productores, consumidores primarios, consumidores de nivel superior y finalmente descomponedores. Estos niveles se utilizan para describir la estructura y dinámica de los ecosistemas. Hay un solo camino a través de una cadena alimentaria. Cada organismo en una cadena alimentaria ocupa un nivel trófico específico (nivel de energía), su posición en la cadena alimentaria o red alimentaria.
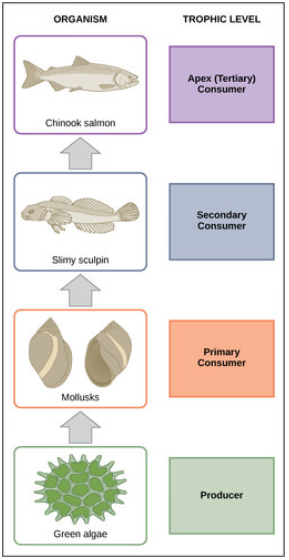
En muchos ecosistemas, la base, o fundamento, de la cadena alimentaria consiste en organismos fotosintéticos (plantas o fitoplancton), a los que se les llama productores. Los organismos que consumen a los productores son herbívoros llamados consumidores primarios. Los consumidores secundarios suelen ser carnívoros que comen a los consumidores primarios. Los consumidores terciarios son carnívoros que comen a otros carnívoros. Los consumidores de nivel superior se alimentan de los siguientes niveles tróficos inferiores, y así sucesivamente, hasta los organismos en la parte superior de la cadena alimentaria. En la cadena alimentaria del lago Ontario, que se muestra en la Figura\(\PageIndex{3}\), el salmón Chinook es el consumidor ápice en la parte superior de esta cadena alimentaria.
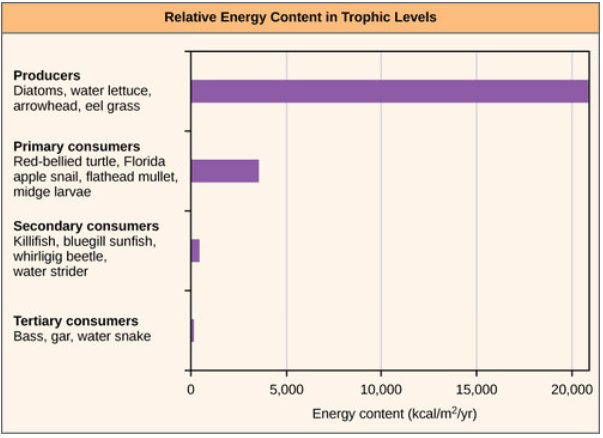
Un factor importante que limita el número de pasos en una cadena alimentaria es la energía. La energía se pierde en cada nivel trófico y entre niveles tróficos como calor y en la transferencia a descomponedores (Figura\(\PageIndex{4}\) a continuación). Por lo tanto, después de un número limitado de transferencias de energía trófica, la cantidad de energía restante en la cadena alimentaria puede no ser lo suficientemente grande como para soportar poblaciones viables en niveles tróficos más altos.
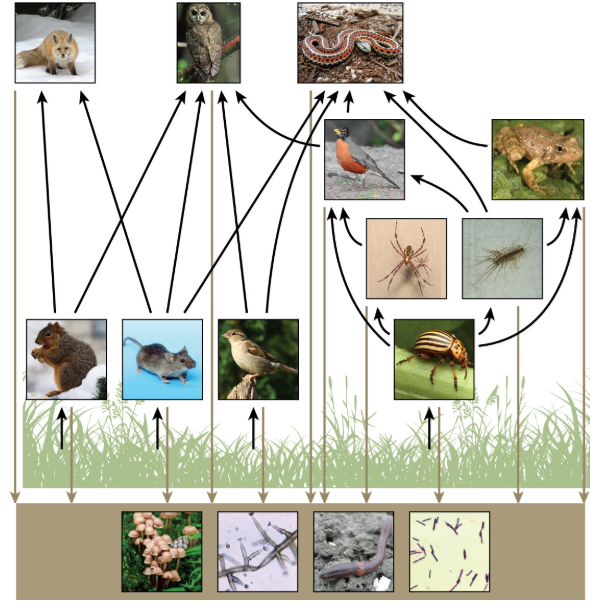
Hay un problema al usar las cadenas alimentarias para describir la mayoría de los ecosistemas. Incluso cuando todos los organismos se agrupan en niveles tróficos apropiados, algunos de estos organismos pueden alimentarse a más de un nivel trófico. Además, las especies se alimentan y son consumidas por más de una especie. En otras palabras, el modelo lineal de ecosistemas, la cadena alimentaria, es una representación hipotética y excesivamente simplista de la estructura de los ecosistemas. Un modelo holístico, que incluye todas las interacciones entre diferentes especies y sus complejas relaciones interconectadas entre sí y con el medio ambiente, es un modelo más preciso y descriptivo para los ecosistemas. Una red alimentaria es un concepto que da cuenta de las múltiples interacciones tróficas (alimentación) entre cada especie (Figura\(\PageIndex{5}\) a continuación).
A menudo se muestran dos tipos generales de redes alimentarias interactuando dentro de un solo ecosistema. Una red alimenticia de pastoreo tiene plantas u otros organismos fotosintéticos en su base, seguidos de herbívoros y diversos carnívoros. Una red alimentaria detrítica consiste en una base de organismos que se alimentan de materia orgánica en descomposición (organismos muertos), incluidos los descomponedores (que descomponen los organismos muertos y en descomposición) y los detritívoros (que consumen detritos orgánicos). Estos organismos suelen ser bacterias, hongos y animales invertebrados que reciclan material orgánico de vuelta a la parte biótica del ecosistema ya que ellos mismos son consumidos por otros organismos.
Cómo los organismos adquieren energía en una red alimentaria
Todos los seres vivos requieren energía de una forma u otra. A nivel celular, la energía se utiliza en la mayoría de las vías metabólicas (generalmente en forma de ATP), especialmente las responsables de construir moléculas grandes a partir de compuestos más pequeños. Los organismos vivos no serían capaces de ensamblar moléculas orgánicas complejas (proteínas, lípidos, ácidos nucleicos e hidratos de carbono) sin un aporte constante de energía.
Los diagramas de la web alimentaria ilustran cómo la energía fluye direccionalmente a través de los ecosistemas. También pueden indicar cuán eficientemente los organismos adquieren energía, la utilizan y cuánto queda para su uso por otros organismos de la red alimentaria. La energía es adquirida por los seres vivos de dos maneras: los autótrofos aprovechan la luz o la energía química y los heterótrofos adquieren energía a través del consumo y digestión de otros organismos vivos o previamente vivos.
Los organismos fotosintéticos y quimiosintéticos son autótrofos, que son organismos capaces de sintetizar su propio alimento (más específicamente, capaces de utilizar carbono inorgánico como fuente de carbono). Los autótrofos fotosintéticos (fotoautótrofos) utilizan la luz solar como fuente de energía, y los autótrofos quimiosintéticos (quimioautótrofos) utilizan moléculas inorgánicas como fuente de energía. Los autótrofos son críticos para los ecosistemas porque ocupan el nivel trófico que contiene productores. Sin estos organismos, la energía no estaría disponible para otros organismos vivos, y la vida no sería posible.
Los fotoautótrofos, como las plantas, las algas y las bacterias fotosintéticas, son la fuente de energía para la mayoría de los ecosistemas del mundo. Los fotoautótrofos aprovechan la energía solar del Sol convirtiéndola en energía química. La tasa a la que los productores fotosintéticos incorporan energía del Sol se denomina productividad primaria bruta. Sin embargo, no toda la energía incorporada por los productores está disponible para los demás organismos en la red alimentaria porque los productores también deben crecer y reproducirse, lo que consume energía. La productividad primaria neta es la energía que permanece en los productores después de contabilizar el metabolismo y la pérdida de calor de estos organismos. La productividad neta está entonces disponible para los consumidores primarios en el siguiente nivel trófico.

Figura\(\PageIndex{6}\). Camarones nadadores, algunas langostas en cuclillas y cientos de mejillones de ventilación se ven en un respiradero hidrotermal en el fondo del océano. Como ninguna luz solar penetra a esta profundidad, el ecosistema está respaldado por bacterias quimioautótrofas y material orgánico que se hunde desde la superficie del océano. Esta foto fue tomada en 2006 en el volcán sumergido NW Eifuku frente a la costa de Japón por la Administración Nacional Oceánica y Atmosférica (NOAA). La cumbre de este volcán altamente activo se encuentra a 1535 m por debajo de la superficie.
Los quimioautótrofos son principalmente bacterias y arqueas que se encuentran en ecosistemas raros donde no se dispone de luz solar, como los asociados con cuevas oscuras o respiraderos hidrotermales en el fondo del océano (Figura\(\PageIndex{6}\)). Muchos quimioautótrofos en respiraderos hidrotermales utilizan sulfuro de hidrógeno (H 2 S), que se libera de los respiraderos, como fuente de energía química. Esto les permite sintetizar moléculas orgánicas complejas, como la glucosa, para su propia energía y, a su vez, abastecer de energía al resto del ecosistema.
Una de las consecuencias más importantes de la dinámica de los ecosistemas en términos de impacto humano es la biomagnificación. La biomagnificación es la concentración creciente de sustancias persistentes y tóxicas en los organismos en cada nivel trófico sucesivo. Se trata de sustancias que son solubles en lípidos y se almacenan en las reservas de grasa de cada organismo. Se ha demostrado que muchas sustancias se biomagnifican, entre ellas estudios clásicos con el pesticida diclorodifeniltricloroetano (DDT), que fueron descritos en el bestseller de la década de 1960 Silent Spring de Rachel Carson. El DDT era un pesticida de uso común antes de que se conocieran sus peligros para los consumidores ápice, como el águila calva. El DDT y otras toxinas son absorbidas por los productores y transmitidas a niveles sucesivos de consumidores a tasas cada vez más altas. Mientras las águilas calvas se alimentan de peces contaminados, sus niveles de DDT aumentan. Se descubrió que el DDT provocó que las cáscaras de huevo de las aves se volvieran frágiles, lo que contribuyó a que el águila calva fuera catalogada como una especie en peligro de extinción según la ley estadounidense. El uso del DDT fue prohibido en Estados Unidos en la década de 1970.
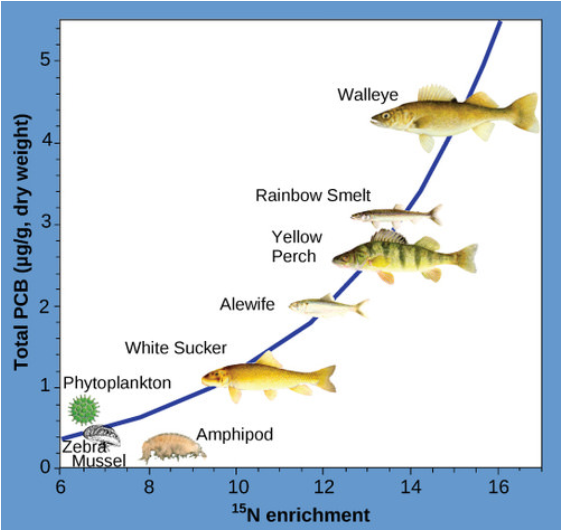
Otra de las sustancias que biomagnifica es el bifenilo policlorado (PCB), el cual se utilizó como líquidos refrigerantes en Estados Unidos hasta que se prohibió su uso en 1979. Los PCB se estudiaron mejor en ecosistemas acuáticos donde las especies de peces depredadores acumularon concentraciones muy altas de la toxina que de otro modo existe a bajas concentraciones en el ambiente. Como se ilustra en un estudio realizado por la NOAA en la Bahía Saginaw del Lago Huron de los Grandes Lagos de América del Norte (Figura\(\PageIndex{7}\) a continuación), las concentraciones de PCB aumentaron desde los productores del ecosistema (fitoplancton) a través de los diferentes niveles tróficos de las especies de peces. El consumidor ápice, el walleye, tiene más de cuatro veces la cantidad de PCB en comparación con el fitoplancton. Además, la investigación encontró que las aves que comen estos peces pueden tener niveles de PCB que son al menos diez veces más altos que los que se encuentran en los peces del lago.
Otras preocupaciones han sido planteadas por la biomagnificación de metales pesados, como el mercurio y el cadmio, en ciertos tipos de mariscos. La Agencia de Protección Ambiental de Estados Unidos recomienda que las mujeres embarazadas y los niños pequeños no consuman ningún pez espada, tiburón, caballa real o tilefish debido a su alto contenido de mercurio. A estos individuos se les aconseja comer pescado bajo en mercurio: salmón, camarón, abadejo y bagre. La biomagnificación es un buen ejemplo de cómo la dinámica de los ecosistemas puede afectar nuestra vida cotidiana, incluso influyendo en los alimentos que comemos.
Lectura suplementaria sugerida
Canales, M. et al. 2018. 6 Cosas que Hacen Posible Como en la Tierra [Infografía]. Geográfico Nacional. Marzo.
Colaboradores y Atribuciones
- Flujo de Energía a través de Ecosistemas de OpenStax está licenciado bajo CC BY 3.0. Modificado del original por Matthew R. Fisher.