
1.8: Técnicas de Campo para Muestreo y Estimación de Poblaciones
- Page ID
- 57787
Las técnicas de campo se refieren a los métodos estandarizados empleados para seleccionar, contar, medir, capturar, marcar y observar individuos muestreados de la población objetivo con el propósito de recolectar los datos requeridos para lograr los objetivos del estudio. El término también incluye métodos utilizados para recolectar especímenes de cupones, muestras de tejido y datos de hábitat. La elección de las técnicas de campo a utilizar para una especie o población en particular está influenciada por cinco factores principales:
- Datos necesarios para lograr los objetivos de inventario y monitoreo
- Alcance espacial y duración del proyecto
- Historia de vida y características de la población
- Terreno y vegetación en el área de estudio
- Restricciones presupuestarias
Requerimientos de Datos
Los tipos de datos requeridos para lograr los objetivos de inventario o monitoreo deben ser la consideración principal en la selección de técnicas de campo. A continuación se discuten cuatro categorías de recolección de datos junto con algunas sugerencias para elegir técnicas de campo adecuadas para cada una.
Datos de ocurrencia y distribución
Para algunos estudios poblacionales, el simple hecho de determinar si una especie está presente en un área es suficiente para realizar el análisis de datos planificado. Por ejemplo, los biólogos que intentan conservar una salamandra amenazada pueden necesitar monitorear el alcance del área de distribución de la especie y el grado de fragmentación de la población en una propiedad de la tierra. Un enfoque hipotético es mapear todos los arroyos en los que se sabe que está presente la salamandra, así como arroyos adicionales que pueden calificar como el tipo de hábitat para la especie en la región. Para monitorear los cambios en la distribución de salamandras, la recolección de datos podría consistir en una encuesta a lo largo de tramos seleccionados aleatoriamente en cada uno de los arroyos para determinar si al menos un individuo (o alguna característica alternativa como la masa de huevo) está presente. Usando solo una lista que incluya el alcance del arroyo (es decir, el identificador único), el año de la encuesta y una variable indicadora de ocupación, un biólogo podría preparar una serie temporal de mapas que mostraran todos los arroyos por año y distinguir el subconjunto de arroyos que se sabía que estaban ocupados por la salamandra. Tal enfoque podría apoyar una evaluación cualitativa de los cambios en el patrón de distribución de especies, logrando así los objetivos del programa, y generar nuevas hipótesis sobre la causa de los cambios observados.
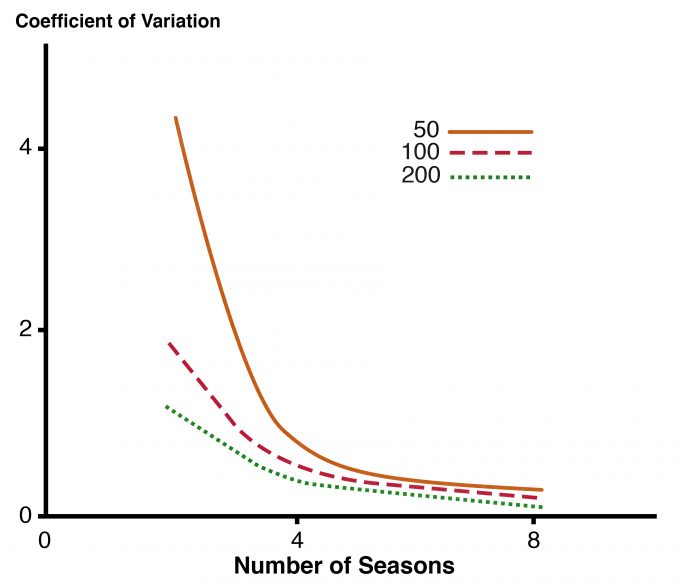
Es mucho más fácil determinar si hay al menos un individuo de las especies objetivo en una unidad de muestreo que contar todos los individuos. Determinar con confianza que una especie no está presente en una unidad de muestreo también requiere un muestreo más intensivo que recolectar datos de conteo o frecuencia porque es muy difícil descartar la posibilidad de que un individuo elude la detección. La probabilidad de ocurrencia se puede estimar utilizando enfoques como los descritos por MacKenzie y Royale (2005). MacKenzie (2005) ofreció una excelente visión general para los gerentes del trade-off entre el número de unidades muestreadas por año y el número de años (u otra unidad de tiempo) para los que se va a realizar el estudio. La variación en la tendencia estimada de ocupación disminuye a medida que aumenta el número de años de recolección de datos (Fig. 8.1). Un nivel similar de precisión se puede lograr mediante la topografía de más unidades durante menos años en comparación con la topografía de menos unidades durante un período más largo.
Tamaño y densidad de la población
La política nacional sobre especies amenazadas y en peligro de extinción se dirige en última instancia a los esfuerzos para aumentar o mantener el número total de individuos de la especie dentro de su área de distribución geográfica natural (Ssuckling y Taylor 2006). El tamaño total de la población y el tamaño efectivo de la población (es decir, el número de individuos reproductores en una población; Lande y Barrowclough 1987) indican más directamente el grado de peligro de especies y la efectividad de las políticas y prácticas de conservación. El tamaño de la población o densidad por unidad de área se suele utilizar como base para los análisis de tendencias, ya que los cambios en la densidad integran cambios en la mortalidad natural, la explotación y la calidad del hábitat. En algunas circunstancias, puede ser factible realizar un censo de todos los individuos de una especie en particular en un área para determinar la densidad poblacional. Normalmente, sin embargo, los parámetros de tamaño y densidad de la población se estiman utilizando análisis estadísticos basados únicamente en una muestra de miembros de la población. Las densidades poblacionales de plantas y animales sésiles se pueden estimar a partir de recuentos tomados en parcelas o datos que describen el espaciamiento entre individuos (es decir, métodos de distancia) y son relativamente sencillas. Los análisis poblacionales de muchas especies animales deben tener en cuenta la respuesta animal a la captura u observación, los sesgos de los observadores y las diferentes probabilidades de detección entre las subpoblaciones. Por lo general, se requieren estudios piloto para recopilar los datos necesarios para abordar estos factores en el análisis. Además, los estudios de marca-recaptura, las encuestas de esfuerzo de captura por unidad y otros métodos de estimación requieren múltiples visitas a unidades de muestreo (Pradel 1996). Estas consideraciones aumentan la complejidad y el costo de los estudios diseñados para la estimación de parámetros poblacionales.
Índices de abundancia
Las metas y objetivos de algunos inventarios biológicos y estudios de monitoreo pueden cumplirse con índices de densidad o abundancia poblacional, más que con estimadores de población. La diferencia entre estimadores e índices es que los primeros producen valores absolutos de densidad poblacional mientras que los segundos proporcionan medidas relativas de densidad que pueden ser utilizadas para comparar índices con poblaciones entre lugares o tiempos. Los índices se basan en el supuesto de que los valores del índice están estrechamente asociados con los valores de un parámetro poblacional, aunque la relación precisa entre el índice y el parámetro generalmente no se cuantifica. Ejemplos de índices de abundancia o densidad son: cobertura de dosel vegetal, número de individuos capturados por 1000 noches de trampa, recuentos de individuos observados durante una unidad de tiempo estandarizada, entre muchos otros. Desde una perspectiva de recolección de datos, los índices de densidad a menudo requieren menos intensidad y complejidad de muestreo que los procedimientos de estimación poblacional Sin embargo, los índices poblacionales no son comparables entre diferentes estudios a menos que las técnicas de campo estén estrictamente estandarizadas Además, rara vez se prueba la suposición de que un índice de abundancia se aproxima estrechamente a la densidad poblacional (Seber 1982).
Datos de aptitud
Para poblaciones raras o en declive, se requieren estimaciones de supervivencia en cada etapa de la vida así como tasas reproductivas. Estos datos no solo proporcionan puntos gatillo útiles para estimar las tasas de declive (lambda) sino que también permiten puntos desencadenantes para la remoción de una especie de un estatus amenazado u otro estatus legal. La recopilación de este tipo de datos suele ser costosa y intensiva en mano de obra. En un estudio sobre búhos manchados del norte, por ejemplo, se han gastado millones de dólares recolectando este tipo de datos (Lint 2001). Esto no es particularmente sorprendente ya que los tipos de datos que serían necesarios para entender la dinámica poblacional de un ave son numerosos y complicados de generar. Las densidades de nidos, los tamaños de nidada, las tasas de eclosión, las tasas de crecimiento y las tasas de supervivencia a la madurez y supervivencia como adultos reproductivos serían un conjunto mínimo de datos. Los nuevos enfoques para estimar las contribuciones individuales al crecimiento poblacional y los cambios en la distribución de rasgos cuantitativos y alelos incluyen análisis genéticos, que pueden conducir a una comprensión aún más detallada del potencial de una población para adaptarse a las variaciones en los factores ambientales (Pelletier et al . 2009).
Estudios de investigación
Los estudios de las relaciones de hábitat o las respuestas de causa y efecto requieren un muestreo coordinado de la población objetivo y mediciones ambientales o factores estresantes a los que la población pueda responder. Los esfuerzos de recolección de datos tienden a ser complejos, requiriendo múltiples protocolos de muestreo para la población objetivo, atributos del sitio de estudio y métricas de patrones de paisaje. El financiamiento requerido para realizar estudios de investigación generalmente limita su aplicación a especies o poblaciones con mayor necesidad de planeación de manejo como las enumeradas como amenazadas o amenazadas. A menudo se llevan a cabo estudios manipuladores para generar los datos necesarios, pero cuando estos se enfocan en una especie amenazada, a menudo surgen preguntas éticas sobre la realización del experimento que ponen a la especie en un riesgo incluso mayor, al menos localmente. De ahí que a menudo es el monitoreo tanto de las condiciones ambientales como de los aspectos de densidad o aptitud poblacional los que se utilizan para evaluar asociaciones en las tendencias entre los parámetros poblacionales y los parámetros ambientales.
Alcance espacial
Claramente, el alcance de inferencia influirá en el tipo de técnica de muestreo utilizada. Las técnicas de atlas de aves reproductoras suelen utilizar grandes cuadrículas colocadas sobre estados enteros para evaluar la ocurrencia de especies en una celda de cuadrícula. Dichos enfoques y los de la Encuesta de aves reproductoras (Sauer et al. 2008) se pueden llevar a cabo a través de esfuerzos voluntarios. Por otro lado, el monitoreo de las tendencias en las tasas reproductivas de búhos moteados del norte, azores del norte o osos grizzly sobre sus rangos geográficos requiere de un presupuesto enorme para recopilar el nivel de datos de población sobre grandes áreas necesarias para comprender las tendencias. Se debe tener mucho cuidado a la hora de decidir qué técnica usar porque tanto los presupuestos como los requisitos de tamaño de la muestra ingresan a la logística. De hecho, a menudo es el equilibrio entre datos más detallados y el costo de producción de esos datos lo que impulsa las decisiones con respecto a los diseños de monitoreo para especies en riesgo.
Técnicas de uso frecuente para el muestreo de animales
La variedad de técnicas disponibles para muestrear animales es amplia y se resume en otros manuales de técnicas (por ejemplo, Bookhout 1994). Resumimos algunos ejemplos de técnicas de uso común, pero sugerimos encarecidamente que aquellos de ustedes que desarrollen planes de monitoreo realicen una búsqueda bibliográfica más completa sobre el muestreo de las especies que son de mayor preocupación en su programa de monitoreo. Primero proporcionamos una breve descripción de las técnicas utilizadas para muestrear vertebrados y luego señalar qué técnicas se utilizan comúnmente entre varios grupos taxonómicos.
Organismos Acuáticos
Algunos organismos acuáticos pueden y han sido monitoreados utilizando técnicas que son esencialmente idénticas a las utilizadas para los vertebrados terrestres. Por ejemplo, en Brasil, los arapaima han sido monitoreados usando una técnica de conteo puntual que cuenta a los individuos a medida que salen a la superficie para la respiración aérea (Castello et al. 2009). Los recuentos de puntos fueron más viables logística y económicamente, se determinaron para representar con mayor precisión los cambios poblacionales a lo largo del tiempo y condujeron a un manejo más efectivo, pero también se intentó una técnica convencional de marca-recaptura con la misma especie de peces (Castello et al. 2009).
Sin embargo, casos como el arapaima son poco frecuentes porque esta especie se detecta al emerger para respiración aérea, tiene una densidad de población lo suficientemente baja en un área lo suficientemente pequeña para ser contada de manera efectiva, y posee ciertas características visuales y acústicas sutiles que permiten la identificación de individuos (Castello et al. 2009). La mayoría de las técnicas utilizadas para muestrear organismos acuáticos son conceptualmente similares a las utilizadas para muestrear organismos terrestres. Pero las limitaciones impuestas a los observadores para tratar el muestreo dentro o sobre el agua y a diversas profundidades del agua requieren que muchas técnicas sean más especializadas. Existe una variedad de técnicas comúnmente utilizadas para muestrear peces y anfibios acuáticos, así como invertebrados acuáticos (Slack et al. 1973). Una evaluación sistemática de alcances de arroyos utilizando encuestas de snorkel (Hankin y Reeves 1988) o equipos de electropesca se usa comúnmente en arroyos y ríos poco profundos (Cunjak et al. 1988).
En estuarios y grandes ríos, los estudios cuantitativos a menudo se confunden por la alta variabilidad de las poblaciones de peces y la alta eficiencia de los aparejos de muestreo de peces (Poizat y Baran 1997). Ante esto, Poizat y Baran (1997) emprendieron un estudio evaluando la eficacia de la encuesta de pescadores en comparación con un enfoque de muestreo de redes de enmalle manejado por científicos y determinaron que combinar ambos enfoques es la mejor manera de aumentar la confianza de que las tendencias observadas son reales. En otras palabras, si ambos conjuntos de datos de encuestas sugieren la misma tendencia, es más seguro inferir que las tendencias son reales que si los conjuntos de datos sugieren tendencias diferentes o si existe solo un tipo de datos.
Las estopas de manta, que son comparables a los métodos de transección de línea pero deben tener en cuenta las condiciones marinas únicas, como la turbidez del agua, las mareas y las características del estado del mar, a menudo se utilizan para monitorear las características generales de los arrecifes de coral y sus poblaciones asociadas. La técnica, que ha sido empleada tanto en programas dirigidos por científicos como en programas comunitarios, consiste en remolcar a un esnorkeler entrenado para observar ciertas variables detrás de una embarcación a velocidad constante a lo largo de un tramo de arrecife determinado (Bass y Miller 1996, Uychiaoco et al. 2005). En un estudio a lo largo de la Gran Barrera de Coral, donde se han empleado remolcadores de manta desde la década de 1970, la línea muestreada se divide en zonas que tardan dos minutos en muestrear y cada dos minutos, la embarcación se detiene para que el observador registre datos en una ficha de datos acuáticos (Bass y Miller 1996). En estas encuestas, los datos suelen incluir recuentos de especies conspicuas, como almejas gigantes, o de ensamblajes enteros, como peces carnívoros y herbívoros, pero la técnica también se utiliza para el monitoreo del hábitat (Bass y Miller 1996, Uychiaoco et al. 2005). De hecho, las observaciones de conjuntos de variables diseñadas para informar a los practicantes sobre el estado de los arrecifes coralinos a lo largo del tiempo, como la pendiente de arrecife, la forma bentónica dominante, el género de coral duro dominante y la complejidad estructural del coral también se registran comúnmente (Bass y Miller 1996, Uychiaoco et al 2005).
Welsh et al. (1987) propusieron un enfoque basado en hábitats para anfibios en pequeñas corrientes de cabecera y también se han utilizado enfoques con limitaciones de tiempo y áreas restringidas para especies de cabecera (Hossack et al. 2006). Las especies reproductoras de estanques o especies que habitan en aguas más profundas a menudo se muestrean usando trampas para pececillos, redes o recuentos de ranas y sapos vocalizantes (Kolozsvary y Swihart 1999, Crouch y Paton 2002).
El seguimiento de animales individuales a través de etiquetas, transpondedores integrados pasivos (etiquetas PIT) y técnicas similares son costosos pero proporcionan información sobre los movimientos de los animales y estimaciones del tamaño de la población y supervivencia. Dichos enfoques han sido utilizados con especies de alto interés como el salmón coho en el noroeste del Pacífico (Wigington et al. 2006).
Organismos Terrestres y Semiacuáticos
La diversidad de formas, tamaños e historias de vida entre los vertebrados terrestres ha llevado al desarrollo de cientos de técnicas de campo diseñadas para diferentes especies y condiciones de sondeo. En el cuadro 8.1 se enumeran las técnicas de campo más utilizadas para la recolección de datos de vida silvestre, pero de ninguna manera es una lista exhaustiva de todos los métodos de inventario y monitoreo. Las técnicas se separan en métodos de observación, captura y marcaje y por el modo por el que se recolectan los datos. Una revisión exhaustiva de todas las diferentes técnicas de campo para organismos terrestres y semiacuáticos es un libro separado en sí mismo (ver Bookhout 1994). Aquí proporcionamos una breve descripción de algunas de las técnicas de uso común.
Kuenzi y Morrison 1998 TagsAves/Mammalsnietfeld et al. 1994
| Modo | Técnica | Grupos objetivo de especies | Referencias | |
| Observacional | Directo | Cuadrados; parcelas de área fija | Organismos sésiles o relativamente inmóviles | Bonham 1989 |
| Recuentos de puntos aviares | Especies de aves que cantan o llaman a territorios | Ralph et al. 1995 | ||
| Mapeo puntual y búsquedas de nidos | Especies de aves territoriales | Ralph et al. 1993 | ||
| Transecto de línea | Grandes mamíferos, aves | Anderson y otros 1979 | ||
| Respuesta de reproducción de llamadas | Lobos, ardillas de tierra, rapaces, pájaros carpinteros | Ogutu y Dublín 1998 | ||
| Búsquedas visuales estandarizadas | Herbívoros grandes, | Cook y Jacobsen 1979 | ||
| Censo | Murciélagos que habitan en las cavernas; grandes | Thomas y Occidente 1989 | ||
| Animal
Signo |
Encuestas de huellas | Mamíferos medio-grandes | Wilson y Delahay 2001 | |
| Recuentos de pellets y scat | Mamíferos medio-grandes | Fuller 1991 | ||
| Búsquedas en caché de alimentos | Grandes carnívoros | Pilcher de Pascuas 1990 | ||
| Estructuras (por ejemplo, guaridas, nidos) | Mamíferos arbóreos; mamíferos fosoriales; osos | Healy y Galés 1992 | ||
| Teledetección | Placas de oruga | Mamíferos medio-grandes | Wilson y Delahay 2001 | |
| Estaciones de foto y video | Mamíferos medio-grandes | Morruzzi et al. 2002 | ||
| Detectores ultrasónicos | murciélagos | Thomas y Occidente 1989 | ||
| Monitoreo de audio | Ranas | Agacharse y Patón 2002 | ||
| Trampas de pelo | Mamíferos pequeños y medianos, grandes carnívoros | McDaniel et al. 2000 | ||
| Telemetría de radio | ¿Limitado por el tamaño corporal del animal (>20 g)? | USGS 1997 | ||
| Telemetría GPS | Limitado por cuerpo animal (>2000 g)? | Girard et al. 2002 | ||
| Radar marino | Murciélagos, aves migratorias | Harmata et al. 1999 | ||
| Radar armónico | Murciélagos, anfibios, reptiles | Pellet et al. 2006 | ||
| Captura | Pasivo | Trampas | Salamandras, lagartos, pequeños mamíferos | Enge 2001, Mengak y Guynn 1987 |
| Trampas Snap | Pequeños mamíferos | Mengak y Guynn 1987 | ||
| Trampas de caja | Mamíferos pequeños y medianos | Powell y Proulx 2003 | ||
| Trampas tipo embudo | Serpientes, tortugas | Enge 2001 | ||
| Fijación de piernas y trampas | Mamíferos grandes | Bookhout 1994 | ||
| Redes de niebla | ||||
| Activo | Unidades a un gabinete | Mamíferos medianos-grandes con respuesta de vuelo | Decalesta y Witmer 1990 | |
| Redes de cañón | Mamíferos medio-grandes | Bookhout 1994 | ||
| Agentes inmovilizadores | Mamíferos grandes | Bookhout 1994 | ||
| Captura de mano | Salamandras | Kolozsvary y Swihart. 1999 | ||
| Marcado | ||||
| Mutilación | Pequeños mamíferos | Madera y Slade 1990 | ||
| Pigmentos | Pequeños mamíferos | Lemen y Freeman 1985 | ||
| Collares y bandas | Aves y mamíferos | Nietfeld et al. 1994 |
Para ciertas especies y condiciones, puede ser factible determinar un recuento de miembros individuales de la población en cuadráticos (parcelas de muestra) colocados de manera aleatoria o sistemática en el área de estudio. Las búsquedas se pueden realizar a pie, vehículos todo terreno o avión dependiendo de la escala y circunstancias de la encuesta. El muestreo cuadrático se usa comúnmente para plantas y elementos de hábitat, pero con animales el muestreo cuadrático plantea algunos desafíos. Si los animales son móviles durante el período de muestreo, entonces debe haber alguna seguridad razonable de que un individuo no se cuenta dos veces en múltiples cuadrantes a medida que se mueve. Se debe considerar el tamaño, el espaciamiento y la movilidad de los organismos.
Los recuentos puntuales son quizás la técnica más utilizada para medir la abundancia y diversidad de aves en bosques templados y pastizales, pero también se han utilizado para estimar la abundancia de otras especies diurnas como las ardillas. Se han descrito variaciones en la técnica para diferentes especies y para satisfacer diferentes necesidades de datos (Verner y Ritter 1985, Verner 1988, Ralph et al. 1995, Huff et al. 2000). Ralph et al. (1995) proporcionaron una colección de artículos que examinan la adecuación del tamaño de la muestra, la detectabilidad de aves, el sesgo de los observadores y las comparaciones entre técnicas.
El mapeo puntual, también conocido como mapeo de territorio, a menudo se utiliza para estimar las densidades de población aviar localizando machos cantantes durante varias visitas al área de estudio y delineando los límites del territorio. La técnica se describe adicionalmente en Ralph et al. (1995). Las búsquedas de nidos se pueden utilizar para evaluar el éxito reproductivo en una población aviar mediante el monitoreo de la supervivencia de huevos y polluelos en el transcurso de una temporada de reproducción. Ambas técnicas son intensivas en mano de obra y no se utilizan comúnmente para inventarios, pero la información obtenida de estos métodos (es decir, densidades territoriales, productividad) puede ser mejores indicadores de tendencias poblacionales y calidad del hábitat que simplemente los recuentos de individuos.
El muestreo de transecto lineal y transecto puntual son métodos de trazado especializados en los que se realiza una búsqueda del organismo diana a lo largo de una franja estrecha que tiene un área conocida. Rara vez se puede suponer que todos los animales son detectados a lo largo del transecto. Sin embargo, si la probabilidad de detección se puede predecir a partir de la distancia entre el animal y la línea central del transecto, entonces se puede usar una función de detección para estimar la densidad de población. El enfoque se puede adaptar a encuestas realizadas a pie, snorkel y vehículos terrestres o aéreos. Buckland et al. (1993) proporcionaron una introducción completa, aunque altamente técnica, a los métodos de transecto lineal y punto transecto. El enfoque ha sido ampliamente aplicado a encuestas de vertebrados, incluyendo tortugas del desierto (Anderson et al. 2001), murrelets veteados (Madsen et al. 1999), pájaros cantores en bosques de roble-pino (Verner y Ritter 1985) y venado mula (White et al. 1989).
Se han utilizado grabaciones de audio de vocalizaciones animales para provocar llamadas y exhibiciones de especies que de otro modo serían difíciles de detectar. La técnica se ha aplicado en estudios de urogallo azul (Stirling y Bendell 1966), búhos moteados del norte (Forsman 1988), ardillas de tierra (Lishak 1977), y otros. El número de respuestas de las especies objetivo provocadas por el registro se calcula durante un intervalo prescrito y proporciona un índice de densidad poblacional.
Las búsquedas visuales estandarizadas se refieren a técnicas utilizadas para determinar la ocurrencia de especies, la riqueza de especies o los valores de densidad relativa, donde el esfuerzo de muestreo se estandariza por espacio o tiempo. Los ejemplos incluyen recuentos de carreteras para mamíferos grandes (Rudran et al. 1996), recuentos de migración de rapaces (Hussell 1981) y encuestas de encuentros visuales para anfibios terrestres (Crump y Scott 1994) (Figura 8.2). Algunas técnicas de búsqueda visual no necesariamente igualan la cantidad de esfuerzo de encuesta entre las unidades de muestreo. En cambio, los recuentos de animales o especies detectadas se estandarizan durante el análisis dividiendo el número de observaciones por una unidad de área o tiempo. La variabilidad entre observadores y condiciones ambientales puede ser fuentes significativas de error no asociadas con la técnica de muestreo y deben evaluarse antes de la recolección de datos para minimizar sesgos y mejorar la precisión.
Bajo ciertas circunstancias, puede ser posible observar de manera efectiva a todos los individuos de la población objetivo. En tales casos, el tamaño de la población puede determinarse directamente a partir del recuento de individuos; no son necesarios procedimientos estadísticos. Los recuentos precisos de los individuos dependen de una tendencia natural de los miembros de la población a agregarse, al menos durante períodos predecibles (por ejemplo, murciélagos que descansan en la cueva) Además, todos los lugares donde se agregan los individuos deben ser conocidos en el área de estudio, y debe haber topógrafos adecuados disponibles para realizar recuentos simultáneos en todas las ubicaciones.
En muchos casos, la población objetivo es altamente críptica o demasiado cautelosa para ser observada directamente y las restricciones presupuestarias impiden que el investigador utilice métodos de captura para la recolección de datos. En estas situaciones, muchas veces es posible inferir la presencia de una especie o determinar un valor índice para la densidad poblacional mediante la observación de signos animales. Los signos son huellas, montones de excrementos, pellets fecales, postes de marcas de aromas o construcciones de animales (por ejemplo, nidos arbóreos, logias de castor, aberturas de madrigueras) que pueden identificarse con precisión como evidencia de una especie en particular. Si las búsquedas de dicha evidencia se realizan en transectos estandarizados o cuadráticos, entonces las observaciones pueden proporcionar un índice confiable de densidad poblacional. Los análisis de datos son similares a los de las observaciones directas. Davis y Winstead (1980) y Wemmer et. al (1996) proporcionan una visión general de los métodos basados en el signo animal.
Las especies esquivas pueden ser muestreadas usando dispositivos de detección remota ubicados en el área de estudio. Las placas de vía (Zelinski y Stauffer 1996) y las trampas para el pelo (Scotts y Craig 1988, McDaniel et al. 2000) son económicas y adecuadas para determinar la ocurrencia y distribución de mamíferos raros en el área de estudio. Los detectores ultrasónicos pueden ser utilizados para monitorear las poblaciones de murciélagos (Kunz et al. 1996), sin embargo no siempre es posible distinguir de manera confiable entre todas las especies de murciélagos. Las estaciones de cámara remota con registradores de datos (Cutler y Swann 1999, Moruzzi et al. 2002) no solo detectan la ocurrencia de la especie, sino que también pueden proporcionar información sobre el sexo, la edad y los patrones de actividad de los individuos. Los cebos en una placa de seguimiento o estación de cámara pueden usarse para aumentar la probabilidad de detectar una especie críptica o rara, pero también pueden sesgar cualquier estimación realizada o atraer desproporcionadamente especies omnívoras comunes, como la zarigüeya de Virginia o el mapache del norte (Figura 8.3).

La radio-telemetría se ha utilizado durante muchos años para recopilar datos sobre la vida silvestre y el movimiento de peces, el tamaño del rango de hogares y la selección del hábitat (Figura 8.4). Los transmisores que pesan <1.0 g ahora están disponibles comercialmente, lo que permite rastrear todos los vertebrados menos los más pequeños. Los sistemas de rastreo que utilizan satélites del sistema de posicionamiento global (GPS) permiten monitorear las ubicaciones de los animales en tiempo real sin requerir que los topógrafos determinen las direcciones de las señales de radio en el campo y son cada vez más razonables. Una colección de resúmenes sobre métodos de telemetría de vida silvestre (USGS 1997) proporciona una introducción útil al tema para estudios terrestres. El radar de banda marina se ha utilizado para contar aves migratorias en puntos de observación (Harmata et al. 1999) y monitorear patrones de actividad de murrelets veteados (Burger 2001).

Finalmente, los análisis genéticos de tejido recolectado de trampas de pelo, scat u otros tejidos han llevado a una explosión de enfoques para evaluar poblaciones, dispersión y patrones evolutivos (Haig 1998, Mills et al. 2000).
Historia de vida y características de la población
Ciertas técnicas son más utilizadas con algunos grupos taxonómicos que con otras. En esta sección te brindamos orientación sobre los tipos de técnicas que podrías considerar dependiendo de las especies incluidas en tu programa de monitoreo.
Anfibios y Reptiles
El pequeño tamaño, la naturaleza críptica y los hábitos fosoriales de las salamandras dificultan especialmente la recolección de datos. Muchos anfibios terrestres se mueven solo distancias cortas y no son susceptibles a técnicas de captura pasiva. Estas especies suelen ser muestreadas mediante búsquedas visuales y técnicas de captura manual con esfuerzo de muestreo estandarizado por área (Jaeger e Inger 1994, Bailey et al. 2004) o una restricción de tiempo (Crump y Scott 1994) (Figura 8.2). Las especies que migran entre estanques reproductivos acuáticos y tipos de hábitat terrestres pueden ser susceptibles a trampas de trampas con cercas de deriva (Corn 1994). Para las especies reproductoras de estanques, las masas de huevos suelen ser más detectables que los adultos de la misma especie, lo que hace que las masas de huevos sean más adecuadas para los estudios Shaffer et al. (1994) y Olson et al. (1997) proporcionaron una excelente introducción a las técnicas para inventarios de anfibios en estanques.
Las encuestas de cobertura han sido ampliamente adoptadas para estimar la abundancia relativa de poblaciones de anfibios y reptiles en diferentes tipos de hábitat (Grant et al. 1992, Harpole y Haas 1999, Engelstoft y Ovaskake 2000). Las tablas de cubierta son objetos como tablas o techos metálicos que proporcionan cobertura diurna a los animales y cuando se levantan revelan una muestra de los animales en la zona que la utilizan (Figura 8.5). De ahí que se trate de una muestra de parcela para aquellas especies que buscan cobertura.
Aves
Como grupo, muchas especies de aves son muestreadas mediante muestreo a distancia porque son muy móviles y vocales (Rostenstock et al. 2002, y ver Buckland et al. 1993 para una visión general del muestreo a distancia). En consecuencia, existen dos medios primarios para detectar muchas especies de aves, aumentando la probabilidad de que puedan ser detectadas, especialmente durante la época de reproducción cuando los machos suelen ser territoriales. Las especies diurnas muestreadas en pastizales, pantanos u otras condiciones de vegetación bastante uniformes a menudo pueden ser muestreadas usando transectos (por ejemplo, Ribic y Muestra 2001). Las muestras tomadas en áreas donde terreno accidentado u otros factores impiden el uso de transectos se basan en recuentos de puntos (Buckland 1987). Otras técnicas de uso común incluyen el mapeo de puntos para comprender las densidades de territorio (por ejemplo, Dobkin y Rich 1998), búsquedas de nidos para comprender las densidades de nidos y variaciones en el método de Mayfield para calcular el éxito de los nidos (Johnson y Shaeffer 1990). Las especies nocturnas presentan desafíos adicionales, pero los recuentos de puntos para los búhos durante su temporada de apareamiento pueden ser efectivos especialmente si se utilizan grabaciones de devolución de llamada para obtener respuestas (Hardy y Morrison 2000). Pero las grabaciones de devolución de llamada pueden introducir sesgos cuando las grabaciones se reproducen en áreas que tienen diferente estructura vegetativa o topografía. Finalmente, las bandas y los retornos de bandas se pueden utilizar para estimar la longevidad y las estructuras de edad de las poblaciones (Pollock y Raveling 1982).
Mamíferos
A menudo se muestrean pequeños mamíferos utilizando trampas de diversas formas (Figura 8.6). Las trampas vivas se utilizan a menudo, ya que son adecuadas para estimaciones de marca-recaptura del tamaño de la población, o las trampas vivas o matadoras se pueden usar para estimar la captura por unidad de esfuerzo estimado de abundancia relativa. Cuando se utiliza cualquier tipo de trampa, se deben revisar y seguir las pautas de bienestar animal.

Las probabilidades de detección están influenciadas por el número de trampas nocturnas (el número de trampas multiplicado por el número de noches muestreadas). Las tasas de captura son fácilmente influenciadas por factores como las interacciones intraespecíficas dependientes de la densidad, el clima y el hábitat, por lo que las probabilidades de captura deben calcularse para permitir una estimación imparcial de la abundancia relativa (Menkens y Anderson 1988). Esto es particularmente importante cuando se evalúan tendencias a lo largo del tiempo durante circunstancias en las que el clima y otras condiciones que afectan a la capturabilidad varían de un año a otro
Los mamíferos más grandes, especialmente aquellos que forman agregaciones sociales o ocurren como poblaciones agrupadas, pueden ser encuestados de manera más efectiva por técnicas de observación que por métodos de captura. Los levantamientos aéreos para ungulados a menudo se realizan utilizando procedimientos de muestreo a distancia, pero nuevamente es necesario considerar la observabilidad entre los tipos de vegetación, las condiciones climáticas y las topografías para asegurar estimaciones imparciales de la abundancia (Pollock y Kendall 1987).
La ocurrencia y los índices de abundancia para los mamíferos medianos a menudo se abordan mediante cámaras activadas remotamente, estaciones de aroma, recuentos de pistas o placas de seguimiento, o encuestas con focos (Figura 8.3; Gese 2001). Cada una de estas técnicas tiene la ventaja de ser razonablemente bajo costo y efectiva en la detección de ciertas especies (ver Gese 2001), pero la estimación del tamaño de la población generalmente no es posible.
Los murciélagos presentan un problema de muestreo único. El uso de registros ultrasónicos de sus llamadas de alimentación puede ayudar a distinguir algunas especies que ocurren en un área (O'Farrell et al. 1999), pero las estimaciones de abundancia a menudo se realizan en sitios de gallinero o maternidad.
Efectos del Terreno y Vegetación
Imagina que estás contando todas las aves que puedas ver u oír a lo largo de un transecto que se extiende desde el centro de un parche boscoso hasta el centro de un campo adyacente. Se desea determinar si la abundancia relativa de aves difiere entre los dos tipos de hábitat. Detectas 28 aves en el bosque y 34 en el campo. ¿Hay más aves en el campo que en el bosque? Quizás. Pero probablemente no. Su capacidad para detectar aves en el bosque se ve obstaculizada por la disminución de la visibilidad en el bosque en comparación con un campo más abierto. De ahí que si corrigieras las diferencias en la detectabilidad en función del tipo de vegetación (utilizando técnicas comunes en el muestreo a distancia) entonces el número de aves detectadas por unidad de área podría ser mucho mayor en el bosque que en el campo. En consecuencia, los recuentos de animales en bruto sin considerar la distancia del observador al animal suelen estar sesgados y no deben usarse como variables de respuesta en los programas de monitoreo. Esto es particularmente importante cuando es probable que la estructura de la vegetación cambie a lo largo del marco de tiempo de monitoreo.
En escalas de tiempo humanas, el terreno no cambia (por lo general) apreciablemente de un periodo de tiempo a otro, pero las observaciones aún deben ser estandarizadas por su detectabilidad en diversos entornos.
Méritos y limitaciones de índices comparados con estimadores
Los índices se utilizan a menudo para evaluar los cambios en las poblaciones a lo largo del tiempo basándose en el supuesto de que algún aspecto de la detección está relacionado con la densidad animal real. Por ejemplo, los siguientes son algunos ejemplos de índices a abundancia:
- recuentos de pistas (Conner et al. 1983)
- Recuentos de grupos de pellets (Fuller 1991)
- tasas de captura (Cole et al. 2000)
- tasas de detección de aves machos cantantes (McGarigal y McComb 1998)
- dominancia relativa (poblaciones de plantas)
- Recuentos de nidos de hoja de ardilla (Healy y Welsh 1992)
- recuentos de logias de castor o cachés (Easter-Pilcher 1990)
Estos ejemplos no proporcionan estimaciones de poblaciones, más bien, se mide un índice con el supuesto de que el índice está relacionado con la población o su aptitud de manera conocida y que los cambios observados en la medición del índice a lo largo del tiempo reflejarán cambios en la población de acuerdo a esta relación. Supuestos como cuantas más pistas se vean, más individuos presentes; o cuantos más pájaros machos se escuchen cantar, más aves se reproducen en el sitio a menudo se hacen. Pero la confiabilidad de estos supuestos se pone en duda, y de hecho la oportunidad de sesgo asociado a los índices a la abundancia es bastante alta. Por ejemplo, los recuentos de huellas pueden estar relacionados con la abundancia animal o con los niveles de actividad animal, pero por otro lado, podrían estar relacionados con las características del sustrato, el clima o cualquier combinación de estas opciones. De igual manera, los recuentos de aves machos cantantes pueden representar tendencias en abundancia de machos territoriales, pero si algunos machos no atraen a una pareja, entonces el número de machos cantantes puede no indicar abundancia de hembras nidificantes ni producción reproductiva. Las tasas de captura de animales en el espacio y el tiempo pueden estar relacionadas con la abundancia animal o con su vulnerabilidad a la captura en diferentes áreas de calidad de hábitat. En consecuencia, aunque a menudo se utilizan índices de abundancia debido a limitaciones logísticas, se debe tener considerable precaución a la hora de interpretar los resultados. En efecto, muchas veces es útil realizar un estudio piloto que permita afirmar con un nivel de certeza conocido cuál es la relación entre el índice y la población real (o aptitud) para la especie que se está monitoreando. Para ello se requiere una estimación de la población.
Los estimadores proporcionan información adicional al usuario y pueden ayudar a abordar algunos de los sesgos inherentes a muchos índices. Afortunadamente hay una serie de herramientas disponibles para estimar abundancias que se basan en la teoría del muestreo y pueden dar como resultado niveles conocidos de confianza colocados alrededor de las estimaciones. Por ejemplo, los estimadores de distancia proporcionan un mecanismo para estimar la abundancia de organismos a partir de puntos o transectos donde la detectabilidad puede diferir entre los tipos vegetativos o los alcances de los arroyos. Los estimadores de población están generalmente disponibles como software libre: https://www.usgs.gov/software/wildlife-software-and-models
Se utiliza una variedad de información como base para estimaciones de poblaciones (captura-recaptura y rendimientos de bandas), algunos de los cuales también pueden proporcionar estimaciones de supervivencia y reproducción. La ventaja abrumadora de usar estimadores de población es que las estimaciones de abundancia, supervivencia y distribución de clases de edad se pueden hacer con estimaciones de confianza. Claramente con muestreo replicado de sitios independientes, los índices a abundancia pueden calcularse con intervalos de confianza, pero aún hay dudas sobre el supuesto de asociación imparcial entre el índice y la población característica de interés. En consecuencia, es importante a la hora de diseñar protocolos de monitoreo para asegurar que se consideren los estimadores, y si no es logísticamente posible utilizar estimadores como variable de respuesta en el programa de monitoreo, entonces el índice seleccionado debe justificarse en relación con su asociación conocida con la población característica de interés. Si esto no se puede hacer, entonces deben señalarse claramente tanto los supuestos como las implicaciones de violar esos supuestos.
Estimación de la estructura comunitaria
Aunque gran parte de este libro se centra en el monitoreo de las especies y las condiciones ambientales en las que viven, en ocasiones los gerentes pueden estar preocupados por mantener o desarrollar condiciones que promuevan comunidades diversas o funcionales, o ser capaces de detectar disminuciones en la diversidad funcional ante estresores ambientales. Las métricas de diversidad han evolucionado a lo largo de décadas y proporcionan un medio para comparar la complejidad entre o entre lugares o tiempos. Desafortunadamente, la mayoría de las métricas de diversidad también entierran información sobre las respuestas de especies individuales dentro de uno o algunos números. Para entender lo que realmente está sucediendo dentro de una comunidad, se debe deconstruir una métrica de diversidad para ver cambios en especies o poblaciones individuales. El diablo efectivamente está en el detalle. Sin embargo, las métricas de diversidad todavía se utilizan como guía para la estructura y función de la comunidad. Por lo general, una métrica de diversidad consiste en una estimación del número de especies en una unidad de espacio y tiempo (riqueza de especies) y la distribución de individuos entre esas especies (uniformidad). En la mayoría de las métricas de uniformidad, el valor máximo alcanzable es 1.00 (un número igual de individuos por unidad de área representado entre todas las especies de la comunidad; es decir, ninguna especie domina la comunidad). Pero consideremos el ejemplo en la tabla 8.2. El hipotético bosque y pastizales tienen la misma diversidad y uniformidad. Entonces, ¿son la misma comunidad? ¿Funcionan de manera similar? Obviamente no. De hecho, el grado en que las dos comunidades son similares en representación de especies y distribución de individuos entre especies en común entre las dos comunidades indica que el porcentaje de similitud entre las dos comunidades es 0. De hecho, cada comunidad tiene más en común con una comunidad más rica en especies y más parejo representada en una sabana que en el bosque o pastizal. Entonces, ¿es mejor la sabana porque es más rica en especies y más diversa? No, es simplemente diferente. En definitiva, estas métricas de estructura comunitaria pueden ser muy difíciles de interpretar sin profundizar en los detalles que comprenden las métricas.
| Comunidad | |||
| Bosque | Savannah | Pastizales | |
| Garbanazo de cabeza negra | 10 | 4 | 0 |
| Gorrión Cantante | 2 | 4 | 0 |
| Robin americano | 4 | 3 | 0 |
| Gorrión saltamontes | 0 | 3 | 4 |
| Reyez de pantano | 0 | 1 | 2 |
| Gorrión de sabana | 0 | 1 | 10 |
| ___ | ___ | ___ | |
| Riqueza de especies | 3 | 6 | 3 |
| Número de individuos | 16 | 16 | 16 |
| La uniformidad de Simpson | 0.797 | 0.956 | 0.797 |
| Diversidad de especies de Simpson 1 | 0.567 | 0.850 | 0.567 |
| Similitud comunitaria (%) 2 | |||
| Forest-Savana | 0.563 | ||
| Bosque-Pastizales | 0.000 | ||
| Sabana-Pastizales | 0.313 | ||
| 1 Blanco (1986) | |||
| 2 Itow (1991) | |||
El muestreo de comunidades de una manera que produzca estimaciones imparciales de la estructura de la comunidad también puede ser problemático. Las evaluaciones de riqueza de especies pueden ser tan simples como elaborar una lista de especies detectadas en un área utilizando técnicas estandarizadas. Pero es probable que algunas especies se detecten con más facilidad que otras. Algunos están activos en diferentes momentos del día, y algunos se mueven más que otros. Por lo que las estimaciones de riqueza se confunden por las diferencias en la detectabilidad. Cuando se calculan métricas de diversidad utilizando estos datos, surgen complicaciones adicionales porque las estimaciones de abundancia para cada especie en la comunidad deben ser imparciales y basadas en la misma unidad de espacio y tiempo. Nuevamente, la detectabilidad diferencial de las especies y los sesgos resultantes del movimiento surgen haciendo que las estimaciones de uniformidad se confundan entre las diferencias reales de abundancia entre especies y las diferencias en la detección entre especies. Las encuestas para determinar la presencia de una especie en un área generalmente requieren menos intensidad de muestreo que el trabajo de campo necesario para recolectar otras estadísticas de población.
Otra métrica que se puede calcular a partir de los datos recopilados dentro de un marco de monitoreo se ocupa de los esfuerzos de muestreo desiguales al intentar estimar el número de especies en un área. Debido a que las especies raras suelen ser menos propensas a ser detectadas que las especies comunes, se puede estimar el número de especies basándose en curvas de rarefacción para que se pueda comparar el número de especies encontradas en dos áreas cuando el esfuerzo de muestreo difirió (Simberloff 1972).
Estimación de la integridad biótica
Karr (1981) desarrolló un índice de integridad biótica (IBI) que fue diseñado para ser utilizado para comparar aspectos de comunidades de peces entre sitios de manera estandarizada que reflejara la calidad del agua del sitio con respecto a su capacidad para apoyar a las comunidades de peces. Incluyó aspectos de la riqueza de especies, la salud de los peces y una serie de otros parámetros. Desde entonces, los investigadores han adaptado esta técnica para otros taxones, incluyendo macroinvertebrados acuáticos y aves (Bryce et al. 2002). Aunque la estructura general de los IBIS es similar a la desarrollada por Karr (1981), cada IBI es típicamente elaborado a una condición de referencia típica de la ecorregión dentro de la cual se produce un área muestreada. De ahí que un IBI desarrollado para el bosque mesofítico mixto de los Apalaches pueda tener una estructura similar a la de uno en las montañas Cascades de Oregón, pero los parámetros medidos se basarían en un conjunto muy diferente de condiciones de referencia. Por esa razón los IBIS no suelen utilizarse en los protocolos de monitoreo terrestre y al igual que los índices de diversidad pueden enterrar información que debe extraerse deconstruyendo el índice.
Revisión del Protocolo de Normalización
Independientemente del enfoque adoptado, ciertos aspectos de un diseño de muestreo deben estandarizarse para minimizar el sesgo en los datos resultantes. Consulte el capítulo 7 para una descripción más detallada de las técnicas de estandarización. Por lo menos se debe prestar atención específica al muestreo consistente de la misma ubicación a lo largo del tiempo, muestreo durante la misma estación y hora del día y utilizando el mismo equipo de un periodo de muestreo al siguiente.
Además de la consistencia en las técnicas de muestreo, ubicaciones y tiempos, la mayoría de los enfoques descritos en este capítulo necesitarán la aprobación de un Comité Institucional de Cuidado y Uso de Animales (IACUC) o junta de revisión comparable en universidades y agencias. Los estándares de cuidado para animales al realizar investigaciones son rigurosos y también deben aplicarse a los programas de monitoreo (ver Laber et al. 2007 y Hafner 2007 para una perspectiva sobre las limitaciones a tales revisiones).
Restricciones presupuestales
La elección de qué técnica de muestreo usar cuando se presentan opciones y un conjunto de metas y objetivos es tan a menudo impulsada por mis limitaciones de tiempo y dinero como por la técnica ideal para adquirir los datos necesarios. Por ejemplo, si bien la información sobre la supervivencia de los juveniles ante el cambio climático puede ser una consideración primordial para una especie de larga vida, un sustituto adecuado podría ser un índice de abundancia de animales jóvenes del año y adultos en la población. Dado que las decisiones presupuestales a menudo se toman con base en la gravedad social del tema, y dado que los valores sociales cambian con el tiempo, también lo hacen los presupuestos. En consecuencia, a menudo es prudente desarrollar un programa de monitoreo utilizando técnicas que sean robustas a las preocupaciones con respecto al sesgo y la precisión y que sean rentables, incluso si no dan como resultado el nivel ideal de datos de estado físico que se necesitarían para responder preguntas demográficas de manera más definitiva. Se podrá recabar información demográfica más detallada de manera complementaria si el nivel de preocupación social llega a un punto en el que se le ordena.
Resumen
Existe una gran variedad de métodos disponibles para detectar, enumerar o evaluar la aptitud de los organismos. Hacer coincidir el enfoque adecuado con las metas y objetivos del estudio es un primer paso clave en el diseño de un programa de monitoreo. Los datos necesarios para lograr los objetivos de inventario y monitoreo suelen ser el factor impulsor para decidir qué técnicas se utilizarán en un programa de monitoreo. Además, la escala del programa de monitoreo también puede influir en los métodos utilizados. Para programas estrechamente enfocados en especies altamente valoradas, las mediciones detalladas de la aptitud pueden ser apropiadas. A medida que aumenta la escala del proyecto sobre el espacio y el tiempo, entonces las estimaciones de abundancia o ocurrencia pueden ser el enfoque más factible.
Además, diferentes taxones varían en su propensión a ser muestreados usando diferentes métodos. Mientras que los animales sésiles pueden ser muestreados mediante muestreo cuadrático, especies de amplio alcance pueden ser muestreadas más fácilmente usando cámaras o trampas de pelo que pueden proporcionar información sobre la ocurrencia o movimientos individuales. De manera similar, simplemente la logística asociada al muestreo en terrenos inhóspitos (para los humanos) limita las elecciones de estrategias de muestreo disponibles. Finalmente, las restricciones presupuestales y logísticas a menudo significan que el sistema de muestreo ideal puede no ser factible porque no se dispone de tiempo o dinero para implementar la técnica para cumplir adecuadamente con las metas y objetivos.
En medio de todas las decisiones con respecto a qué técnica emplear, debe estar continuamente consciente de la necesidad de estandarizar enfoques en el espacio y el tiempo y buscar minimizar cualquier sesgo en sus métodos de muestreo.
Referencias
Anderson, D.R., J.L. Laake, B.R.Cran, y K.P. Burnham. 1979. Lineamientos para el muestreo transecto lineal de poblaciones biológicas. Diario de Manejo de Vida Silvestre 43:70—78.
Anderson, D.R., K.P. Burnham, B.C. Lubow, L. Thomas, P.S. Maíz, P.A. Medica, y R.W. Marlow. 2001. Ensayos de campo de los métodos de transecto lineal aplicados a la estimación de la abundancia de tortugas del desierto. Diario de Manejo de Vida Silvestre 65:583—597.
Bailey, L.L., T.R.Simons, y K.H. Pollock. 2004. Estimación de la ocupación del sitio y los parámetros de probabilidad de detección de especies para salamandras terrestres. Aplicaciones Ecológicas 14:692 —702.
Bajo, D.K. e I.R. Miller. 1996. Encuestas de estrellas de mar y corales de corona de espinas utilizando las técnicas de rastreo de manta y búsqueda de buceo. Monitoreo a largo plazo del procedimiento operativo estándar de la Gran Barrera de Coral número 1. Instituto Australiano de Ciencias Marinas, Townsville.
Bonham, C. D. 1989. Mediciones de vegetación terrestre. John Wiley, Nueva York, Nueva York, EUA. 338pp.
Bookhout, T.A. (ed). 1994. Técnicas de investigación y manejo para vida silvestre y hábitats. La Sociedad de Vida Silvestre, Bethesda, Maryland. 740 pp.
Bryce, S.A., R.M. Hughes, y P.R. Kaufman. 2002. Desarrollo de un índice de integridad de aves: utilizando conjuntos de aves como indicadores de condición ribereña. Gestión Ambiental 30:294 —310.
Buckland, S.T. 1987. Sobre el método de parcela circular variable para estimar la densidad animal. Biometría 43:363 —384.
Buckland, S.T., D.R. Anderson, K P. Burnham, y J.L. Laake. 1993. Muestreo a distancia: Estimación de la abundancia de poblaciones biológicas. Chapman y Hall, Nueva York. 446pp.
Hamburguesa, A.E. 2001. Uso de radar para estimar poblaciones y evaluar asociaciones de hábitats de Murrelets veteados. Diario de Manejo de Vida Silvestre 65:696—715.
Castello, L., J.P. Viana, G. Watkins, M. Pinedo-Vásquez, y V.A. Luzadis. 2009. Lecciones de la integración de pescadores de Arapaima en el manejo de la pesca en pequeña escala en la Reserva Mamirauá, Amazonía. Gestión Ambiental 43:197-209.
Cole, E.C., W.C. McComb, M, Newton, J.P. Leeming, y C.L. Chambers. 1998. Respuesta de pequeños mamíferos a la tala, quema y aplicación de glifosato en la cordillera de la costa de Oregón. Revista de Manejo de Vida Silvestre 62:1207 —1216.
Conner, M.C., R.F. Labisky, y D.R. Progulske, Jr. 1983. Índices de estación de olor como medidas de abundancia poblacional para linces, mapaches, zorros grises y zarigüeyas. Boletín de la Sociedad de Vida Silvestre 11, 146—152.
Cook, R.D., y J.O. Jacobson. 1979. Un diseño para estimar el sesgo de visibilidad en levantamientos aéreos. Biometría 35:735 —742.
Maíz, P.S. 1994. Cercas de deriva en línea recta y trampas de trampas. Páginas 109-117 En W.R. Heyer, M A. Donnelly, R.W. McDiarmid, L.C. Hayek, y M.S. Foster, editores. Medición Y Monitoreo de la Diversidad Biológica: Métodos Estándar para Anfibios,. Instituto Smithsoniano. Prensa, Washington, D.C.
Crouch, W.B., y P.W.C. Paton. 2002. Evaluar el uso de encuestas de llamadas para monitorear anuros reproductivos en Rhode Island. Revista de Herpetología 36, 185—192.
Crump, M.L., y N.J. Scott Jr. 1994. Encuestas de encuentros visuales. Páginas 84—92 En W.R. Heyer, M.A. Donnelly, R.W. McDiarmid, L.C. Hayek, y M.S. Foster, editores. Medición y Monitoreo de la Diversidad Biológica: Métodos Estándar para Anfibios,. Smithsonian Institution Press, Washington, DC.
Cunjak, R.A., R.G. Randall, y E.M.P. Chadwick. 1989. Snorkeling versus electropesca: una comparación de técnicas censales en ríos salmónidos del Atlántico. Naturaliste Canadien. 115:89-93.
Cutler, T., y D.E. Swann. 1999. Uso de la fotografía remota en la ecología de vida silvestre: una revisión. Wildl. Soc. Bol. 27:571-581.
Davis, D.E. y R.L. Winstead,. 1980. Estimación del número de poblaciones de vida silvestre. Páginas. 221—245.En S.D. Schemnitz, editor, Manual de técnicas de manejo de vida silvestre, The Wildlife Society, Boston.
Decalesta, D.S. y G.W. Witmer. 1990. Censo drive-line para venados dentro de recintos cercados. USDA Papel de Investigación del Servicio Forestal NE-643. 4pp.
Dobkin D.S., y A.C. Rich. 1998. Comparación de line-transect, spotmap y encuestas puntuales para aves en áreas ribereñas de la Gran Cuenca. Diario de Ornitolología de Campo 69:430 —443
Pascua-Pilcher, A. 1990. Tamaño de caché como índice al tamaño de colonia de castor en el noroeste de Montana. Boletín de la Sociedad de Vida Silvestre 18:110 —113.
Elzinga, C.L., D.W. Salzer y J.W.Willoughby. 1998. Medición y monitoreo de poblaciones vegetales. Bureau of Land Management, Referencia Técnica 1730—1, Denver, Colorado, EUA.
Enge, K.M. 2001. Los escollos de las trampas de trampas. Revista de Herpetología 35:467 —478.
Engelstoft, C. y K.E. Ovaskake. 2000. Objetos de cobertura artificial como método de muestreo de serpientes (Contia tenuis a nd Thamnophis spp.) en Columbia Británica. Naturalista del Noroeste 81:35 —43.
Forsman, E.D. 1988. Un estudio de búhos manchados en bosques jóvenes en la cordillera costera norte de Oregón. Murrelet 69:65-68.
Fuller, T.K. 1991. ¿Los recuentos de pellets indexan los números de venados cola blanca y cambian la población Revista de Manejo de Vida Silvestre 55, 393—396.
Gese, E.M., 2001. Monitoreo de poblaciones de carnívoros terrestres. Páginas 373—396 En: J.L. Gittleman et al., editores. Conservación de Carnívoros. Cambridge University Press, Cambridge, Reino Unido.
Girard, I., J.P. Ouellet, R. Courtois, C. Dussault, y L. Breton. 2002. Efectos del esfuerzo de muestreo basado en telemetría GPS en estimaciones de tamaño de rango doméstico. Diario de Manejo de Vida Silvestre 66:1290 —1300.
Grant, B.W., A.D. Tucker, J.E. Lovich, A.T. Mills, P.M. Dixon, y J.W. Gibbons. 1992. El uso de cobertores en la estimación de patrones de abundancia de reptiles y anfibios. Páginas 379—403 En R. Seigel y N. Scott, editores. Vida silvestre 2001, Elsevier Science Publ., Inc., Londres, Reino Unido.
Haig, S. 1998. Aportes moleculares a la conservación. Ecología 79:413 —425.
Hafner, M.S. 2007. La investigación de campo en la mamología: una empresa en peligro. Diario de Mammalogía 88:1119 —1128.
Hankin, D.G., y G.H. Reeves. 1988. Estimación de la abundancia total de peces y el área total del hábitat en pequeños arroyos con base en métodos de estimación visual. Revista Canadiense de Pesca y Ciencias Acuáticas 45:834 —844.
Hardy, P.C. y M.L. Morrison. 2000. Factores que afectan la detección de búhos elfos y búhos chirridos occidentales. Boletín de la Sociedad de Vida Silvestre 28:333 —343.
Harmata, A.R., K.M. Podruzny, J.R. Zelenak, y M.L. Morrison. 1999. Uso de radar de vigilancia marina para estudiar los movimientos de aves y la evaluación de impacto. Boletín de la Sociedad de Vida Silvestre 27:44 —52.
Harpole, D.N. y C.A. Haas. 1999. Efectos de siete tratamientos silvícolas en salamandras terrestres. Ecología y Manejo Forestal 114:349-356.
Healy, W.M. y C.J.E. Welsh. 1992. Evaluación de transectos de línea para monitorear poblaciones de ardillas grises. Boletín de la Sociedad de Vida Silvestre 20:83 —90.
Hossack, B.R., P.S. Maíz, y D.B. Fagre. 2006. Patrones divergentes de abundancia y estructura de clase de edad de renacuajos de corriente de cabeceras en cuencas quemadas y no quemadas. Revista Canadiense de Zoología 84:1482 —1488.
Huff, M.H., K.A. Bettinger, H.L. Ferguson, M.J. Brown, y B. Altman. 2000. Un protocolo de conteo puntual basado en hábitats para aves terrestres, enfatizando Washington y Oregón. Informe Técnico General del Servicio Forestal de Estados Unidos PNW-GTR-501
Hussell, D.J.T. 1981. El uso de recuentos migratorios para el monitoreo de los niveles de población de aves. Páginas 92—102 en C. J. Ralph y J. M. Scott, editores. Estimación de Números de Aves Terrestres. Estudios en Biología Aviar, núm. 6.
Iow, S. 1991. Rotación de especies y patrones de diversidad a lo largo de una coenoclina de bosque de hoja ancha de hoja perenne. J. Veg. Sci. 2:477-484.
Jaeger, R.G., y R.F.Iinger. 1994. Muestreo cuadrático. pp. 97—102 En W.R. Heyer, M.A. Donnelly, R.W. McDiarmid, L.C. Hayek, y M.S. Foster, editores. Medición y monitoreo de la diversidad biológica: Métodos estándar para anfibios, Smithsonian Institution Press, Washington, DC.
Johnson, D.H., y T.L. Shaffer. 1990. Estimando el éxito del nido: cuando gana Mayfield. Auk 107:595—600.
Karr, J.R. 1981. Evaluación de la integridad biótica utilizando comunidades de peces. Pesquerías 6: (6) 21—27.
Kolozsvary M.B., y R.K. Swihart. 1999. Fragmentación del hábitat y distribución de anfibios: correlatos de parches y paisajes en tierras de cultivo. Revista Canadiense de Zoología 77:1288 —1299
Kuenzi A.J. y M.L. Morrisson. 1998. Detección de murciélagos mediante redes de niebla y sensores ultrasónicos. Boletín de la Sociedad de Vida Silvestre 26:307-311.
Kunz, T.H., D.W. Thomas, G.C. Richards, C.R. Tidemann, E.D. Pierson, y P.A.
Racey. 1996. Técnicas de observación para murciélagos. Páginas 105—114 En: D.E. Wilson, F.R. Cole, J.D. Nichols, R. Rudran, y M.S. Foster, Editores. Medición y Monitoreo de la Diversidad Biológica. Smithsonian Institution Press, Washington, D.C.
Laber, K., B.W. Kennedy, y L. Young. 2007. Estudios de campo y el IACUC: revisión de protocolos, supervisión y consideraciones de salud y seguridad ocupacional. Animal de Laboratorio 36:27 —33.
Lande, R. y G.F. Barrowclough. 1987. Tamaño efectivo de la población, variación genética y su uso en el manejo poblacional. Páginas 87-123 en: M.E. Soulé, Editor, Poblaciones viables para la conservación, Cambridge University Press. Cambridge, Reino Unido. 204pp.
Leman, C.A., y P.W. Freeman. 1985. Seguimiento de mamíferos con pigmentos fluorescentes: una nueva técnica. Diario de Mammalogía 66:134 —136
Lint, J. 2001. Plan de monitoreo de efectividad del búho manchado del norte bajo el Plan Forestal Noroeste: Informe resumido anual 2000 Programa de Monitoreo Interagencial del Plan Forestal Noroeste, Oficina Regional de Ecosistemas, Portland, OR.
Lishak, R. S. 1977. Censando ardilla de tierra de 13 líneas con llamadas de alarma para adultos y jóvenes. Revista de Manejo de Vida Silvestre 41:755 —759.
MacKenzie, D.I. 2005. ¿Estaba ahí? Tratar la detección imperfecta de datos de presencia/ausencia de especies. Australia y Nueva Zelanda Journal of Statistics 47:65 —74.
MacKenzie, D.I., J.D. Nichols, J.E. Hines, M.G. Knutson, y A.D. Franklin. 2003. Estimar la ocupación del sitio, la colonización y las probabilidades de extinción local cuando no se detecta una especie con certeza. Ecología 84:2200 —2207.
MacKenzie, D.I., y J.A. Royle. 2005. Diseñar estudios de ocupación eficientes: asesoría general y consejos sobre asignación del esfuerzo de encuesta. Revista de Ecología Aplicada 42:1105 —1114.
Madsen, S., D. Evans, T. Hamer, P. Henson, S. Miller, S.K. Nelson, D. Roby, y M Stapanian. 1999. Plan de monitoreo de efectividad de murrelet veteado para el Plan Forestal Noroeste. Servicio Forestal del USDA Informe Técnico General PNW-GTR-439. 51 pp.
McDaniel, G.W., K.S. McKelvey, J.R. Squires, y L.F. Ruggiero. 2000. Eficacia de señuelos y trampas capilares para detectar lince. Boletín de la Sociedad de Vida Silvestre 28:119 —123.
McGarigal, K. y W. C. McComb. 1999. Fragmentación forestal y comunidades de aves reproductoras en la Cordillera de Oregón. Capítulo 13 en J. P. Rochelle, L. A. Lehman, y J. Wisniewski, editores. Fragmentación forestal: Vida Silvestre e Implicaciones de Manejo Brill Press, Países Bajos.
Mengak, M.T., y D.C. Guynn. 1987. Trampas y trampas para muestreo de pequeños mamíferos y herpetofauna. American Midland Naturalista 118:284 —288.
Menkens, G.E., y S.H. Anderson. 1988. Estimación del tamaño de la población de pequeños mamíferos. Ecología 69:1952 —1959.
Mills L.S., J.J. Citta, K.P. Lair, M.K. Schwartz, y D.A. Tallmon. 2000. Estimación de la abundancia animal mediante muestreo de ADN no invasivo: promesa y escollos. Aplicaciones Ecológicas 10:283 —294.
Moruzzi, T.L., T.K. Fuller, R.M. DeGraaf, R.T. Brooks, y W.J. Li. 2002. Evaluación de cámaras activadas remotamente para topografía de distribución de carnívoros. Boletín de Sociedad de Vida Silvestre 30:380-386
Nichols, J.D., J.E. Hines, J.R. Sauer, F.W. Fallon, J E. Fallon, y P.J. Heglund. 2000. Un enfoque de doble observador para estimar la probabilidad de detección y abundancia a partir de recuentos puntuales. Auk 117:393 —408.
Nietfeld, M. T., M. W. Barrett, y N. Silvey. 1994. Técnicas de marcado de vida silvestre. Páginas 140-168 En T. A. Bookhout, editor. Técnicas de investigación y manejo para vida silvestre y hábitats. La Sociedad de Vida Silvestre, Bethesda, Md.
O'Farrell, M.J., B.W. Miller, y W.L. Gannno.1999. Identificación cualitativa de murciélagos de vuelo libre utilizando el detector Anabat. Diario de Mammalogía 80:11 —23.
Ogutu, J.O. y H.T. Dublin. 1998. La respuesta de leones e hiaenas manchadas a las retransmisiones sonoras como técnica para estimar el tamaño de la población. Revista Africana de Ecología 36:83—95.
Olson, D.H., W.P. Leonard, y R.B. Bury. 1997. Muestreo de anfibios en hábitats lénticos: métodos y enfoques para el noroeste del Pacífico. Sociedad de Biología de Vertebrados Noroeste, Olimpia, Lavado.
Pellet, J., L. Rechsteiner, A.K. Skrivervik, J.F. Zürcher, y N. Perrin. 2006. Uso del buscador de dirección armónica para estudiar los hábitats terrestres de la rana arbórea europea (Hyla arborea). Anfibia-Reptilia 27:138-142.
Pelletier, F., D. Reale, J. Watters, E. H. Boakes, y D. Garant. 2009. Valor de las poblaciones cautivas para la investigación genética cuantitativa. Tendencias en Ecología y Evolución 24:263-270.
Poizat, B. y E. Baran. 1997. El conocimiento de los pescadores como antecedentes en la ecología de peces tropicales: una comparación cuantitativa con los resultados de muestreo de peces. Biología Ambiental de los Peces. 50 (4) :435-449.
Pollock, K.H., y D.G. Raveling. 1982. Supuestos de modelos modernos de recuperación de banda, con énfasis en tasas de supervivencia heterogéneas. Revista de Manejo de Vida Silvestre 46:88 —98.
Pollock, K.H., y W.L. Kendall. 1987. Visibilidad en levantamientos aéreos: Una revisión de los procedimientos de estimación. Revista de Manejo de Vida Silvestre 51:502 —510.
Powell, R.A. y G. Proulx. 2003. Atrapar y marcar mamíferos terrestres para investigación: integrando ética, criterios de desempeño, técnicas y sentido común. Diario Ilar 44:259 —276.
Pradel, R. 1996. Utilización de captura-marca-recaptura para el estudio de las tasas de reclutamiento y crecimiento poblacional. Biometría 52 703-709.
Ralph, C.J., S. Droege, y J.R. Sauer. 1995. Manejo y monitoreo de aves mediante recuentos puntuales: Normas y aplicaciones. Páginas 161—168 en C. J. Ralph, J. R. Sauer, y S. Droege, Editores. Monitoreo de Poblaciones de Aves por Recuentos de Puntos.Servicio Forestal del USDA Informe Técnico General PSW-GTR-149.
Ralph, C.J., G.R. Geupel, P. Pyle, T.E. Martin, y D.F. DeSante. 1993. Manual de métodos de campo para el monitoreo de aves terrestres. Servicio Forestal del USDA Informe Técnico General PSW-GTR-144. 41pp.
Ribic, C.A., y D.W. Muestra. 2001. Asociaciones de aves de pastizales con factores paisajísticos en el sur de Wisconsin. American Midland Naturalista 146:105 —121.
Rosenstock, S.S., D.R. Anderson, K.M. Giesen, T. Leukering, y M.F. Carter. 2002. Técnicas de conteo de aves terrestres: Prácticas actuales y una alternativa. Auk 119:46 —53.
Rudran, R, T.H. Kunz, S.C. Jarman, y A.P. Smith. 1996. Técnicas observacionales para mamíferos no volantes. Páginas 81—114 En D.E. Wilson, F.R. Cole, J.D. Nichols, R. Rudran, y M.S. Foster, Editores. Medición y monitoreo de la diversidad biológica. Métodos estándar para mamíferos. Institución Smithsonian, Washington, DC, EUA.
Sauer, J.R., J.E. Hines, y J. Fallon. 2008. La Encuesta, Resultados y Análisis de Aves Creadoras en América del Norte 1966 — 2007. Versión 5.15.2008. USGS Patuxent Wildlife Research Center, Laurel, MD
Scotts, D.J., y S.A. Craig, S.A. 1988. Tubo de muestreo capilar mejorado para la detección de mamíferos raros. Australian Wildlife Research 15:469 —472.
Seber, G.A.F. 1982. La estimación de la abundancia animal, 2a ed. Griffin Publishers, Londres. 654pp.
Shaffer H.B., R.A. Alford, B.D. Woodward, S.J. Richards, R.G. Altig, y C. Gascón. 1994. Muestreo cuantitativo de larvas de anfibios. Páginas 130—141 En: Heyer W.R., M.A. Donnelly, R.W. McDiarmid, L.C. Hayek, y M.S. Foster., editores. Medición y Monitoreo de la Diversidad Biológica: Métodos Estándar para Anfibios. Smithsonian Institute Press, Washington, D.C.
Simberloff, D.S. 1972. Propiedades de la medición de la diversidad de rarefacción. Naturalista Americano 106:414-418.
Simberloff, D.S. 1978. Uso de rarefacción y métodos afines en ecología. pp. 150-165 En K.L. Dickson, J. Cairns, Jr., y R.J. Livingston, Editores. Datos biológicos en la evaluación de la contaminación del agua: análisis cuantitativo y estadístico. Sociedad Americana de Pruebas y Materiales STP 652, Filadelfia, PA.
Slack, K.V., R.C. Averett, P.E. Greeson, y R.G. Lipscomb. 1973. Métodos de recolección y análisis de muestras biológicas y microbiológicas acuáticas: Técnicas del Servicio Geológico de los Estados Unidos de Investigaciones de Recursos Hídricos. Número de Reporte 05-A4.
Stirling, D. F, y J. F. Bendell. 1966. Censo de urogallo azul con llamadas grabadas de la hembra. Revista de Manejo de Vida Silvestre 30:184 —187.
Sucking, K. y M. Taylor. 2006. Hábitat crítico y recuperación. Páginas 50-67 en D.D. Goble, J.M. Scott y F.W. Davis. La Ley de Especies Amenazadas a los 30: Vol. 1: Renovando la Promesa de Conservación. Island Press. Washington, D.C.
Thomas, D.W., y S.D. West. 1989. Métodos de muestreo para murciélagos. USDA Servicio Forestal Informe Técnico General PNW 243. 20pp.
Servicio Geológico de Estados Unidos. 1997. Foro sobre telemetría de vida silvestre: innovaciones, evaluaciones y necesidades de investigación; 21-23 de septiembre de 1997, Snowmass Village, Colorado. Programa y Resúmenes. Servicio Geológico de Estados Unidos y The Wildlife Society.
Uychiaoco, A.J., H.O. Arceo, S.J. Green, M.T. De la Cruz, P.A. Gaite, y P.M. Ali ño. 2005. Monitoreo y evaluación de áreas protegidas de arrecifes por pescadores locales en Filipinas: endurecimiento del ciclo de manejo adaptativo. Biodiversidad y Conservación 14:2775-2794.
Verner, J., y L.V. Ritter. 1985. Comparación de transectos y recuentos puntuales en bosques de roble y pino de California. Cóndor 87:47 —68.
Verner, J. 1988. Optimización de la duración de los recuentos puntuales para monitorear tendencias en poblaciones de aves. Nota de Investigación del Servicio Forestal del USDA PSW-395.
Galés, H.H., Jr. 1987. Monitoreo de herpetofauna en hábitats boscosos del noroeste de California y suroeste de Oregón: una estrategia integral. Páginas 203-213 en T.R Plumb y N.H. Pillsbury, editores. Manejo de múltiples usos de los recursos de madera dura de California. USDA Servicio Forestal Informe Técnico General PSW-100.
Wemmer C., T.H. Kunz, G. Lundie-Jenkins, y W. McShea. 1996. Signo de mamíferos. Páginas 157-176 En D.E. Wilson, F.R. Cole, J.D. Nichols, R. Rudran, y M.S. Foster Editores. Medición y Monitoreo de la Diversidad Biológica — Métodos Estándar para Mamíferos. Smithsonian Institution Press, Washington, DC.
White, G.C., R.M. Bartmann, L.H. Carpenter, y R.A. Garrott. 1989. Evaluación de transectos de línea aérea para estimar densidades de ciervos mulos. Revista de Manejo de Vida Silvestre 53:625 —635.
Blanco, M.J. 1986. Medidas de segregación y diversidad en la distribución poblacional. Índice de Población 52:198 —221.
Wigington, P.J., J.L. Ebersole, M.E. Colvin, S.G. Leibowitz, B. Miller, B. Hansen, H. Lavigne, D. White, J.P. Baker, M.R. Church, J.R. Brooks, M.A. Cairns, y J.E. Compton. 2006. Dependencia del salmón coho en arroyos intermitentes. Fronteras en Ecología y Medio Ambiente 4:513-518.
Wilson, G.J. y R.J. Delahay. 2001. Revisión de métodos para estimar la abundancia de carnívoros terrestres utilizando señales de campo y observación. Investigación de Vida Silvestre 28:151 —164.
Wood, M.D. y N.A. Slade. 1990. Comparación de marcaje de orejas y recorte de dedos en topillos de pradera, Microtus ocrogaster. Diario de Mammalogía 71:252—255.
Zielinski, W.J. y H.B. Stauffer. 1996. Monitoreo de poblaciones de Martes en California: diseño de encuestas y análisis de potencia. Aplicaciones Ecológicas 6:1254—1267.