
1.13: Usos de los Datos- Síntesis, Evaluación de Riesgos y Toma de Decisiones
- Page ID
- 57786
Imagina el siguiente escenario. Acaba de gastar casi 500.000 dólares en los últimos cinco años recopilando información sobre los cambios en la abundancia de serpientes de cola afilada en las estribaciones del valle de Willamette en Oregón. Se recolectaron datos de 30 sitios seleccionados al azar en terrenos públicos que lograron restaurar sabanas de encino de Oregón, y en otros 30 sitios en tierras privadas que son pastoreadas. Los datos se presentan en la figura 13.1.
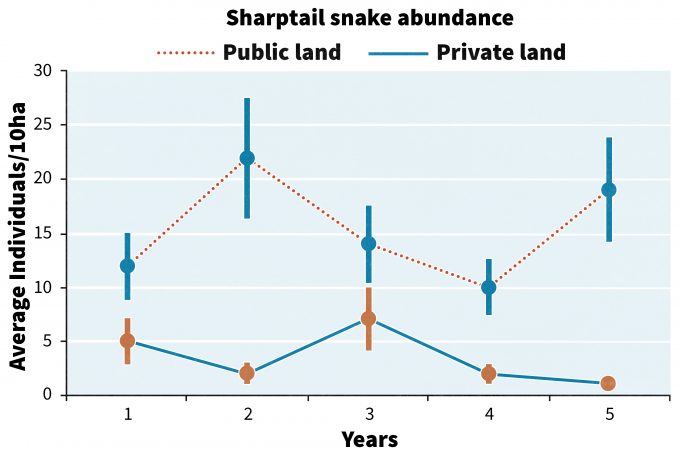
Entonces, dada esta información, ¿qué haces? ¿Continuar monitoreando? ¿Utiliza la información para realizar cambios? ¿Cuáles son los riesgos de cambiar vs. continuar con el status quo? ¿Se pueden integrar estos datos con datos de monitoreo de otros programas para crear una imagen más amplia del estado de los ecosistemas de Oregón? Seguiremos este ejemplo a través de algunos pasos clave en la interpretación de los datos de monitoreo y veremos cómo se podrían tomar las decisiones.
Umbrales y puntos de activación
Claramente hay una serie de temas que deben ser considerados no sólo por los directivos sino también por los stakeholders antes de realizar cualquier cambio. Un enfoque es acordar con las partes interesadas al inicio del programa de monitoreo que si se alcanza un determinado umbral o punto de activación, entonces se implementarán acciones alternativas de manejo. Block et al. (2001) diferenciaron entre puntos gatillo que inician un cambio para promulgar la recuperación, y umbrales, que indican éxito en una acción de recuperación. En el caso de la figura 13.1, un punto de activación puede estar registrando < 5 serpientes/10ha por dos o más años consecutivos. Si se llega a tal punto de activación, se podría acordar previamente con los actores interesados que el organismo responsable tomaría una serie de pasos para restaurar el hábitat de la especie. O en el caso de especies en peligro de extinción, se podrá tomar la decisión de capturar individuos e iniciar un programa de cría en cautividad. Pero en nuestro caso hipotético después de 5 años, las detecciones de serpientes de cola afilada cumplen con el punto desencadenante en el año 5, por lo que en ese momento los biólogos de la agencia de gestión pública pueden comenzar a reunirse con propietarios privados para explorar las siguientes opciones para restaurar el hábitat de la especie:
- Brindar asistencia a los propietarios en la restauración del hábitat
- Ofrecer incentivos a los propietarios para que alteren el pastoreo y otras prácticas de uso de la tierra.
- Explore la compra de una servidumbre de conservación que permita a los biólogos públicos administrar tierras
- Explore la compra de propiedades clave y comience la restauración del hábitat
Cualquiera de las opciones anteriores puede ser aceptable para un terrateniente pero no para otro. A medida que se implementan estas u otras opciones, el monitoreo continuo puede permitir la detección del punto en el que se supera y mantiene un umbral de recuperación, digamos >10 serpientes/10 ha por >2 años. El monitoreo de un área de control para comprender los cambios en la abundancia en las tierras públicas de conservación proporcionará un punto de comparación para ayudar a garantizar que los patrones vistos en tierras privadas utilizando los enfoques anteriores sean más probables causados por acciones de manejo que por otros efectos extraños. Por ejemplo, si la abundancia tanto en las tierras públicas como privadas disminuyó con el tiempo a pesar de los cambios en las prácticas de manejo en las tierras privadas, entonces las disminuciones son más probables debido a factores no asociados con el manejo como los cambios en el clima o las enfermedades.
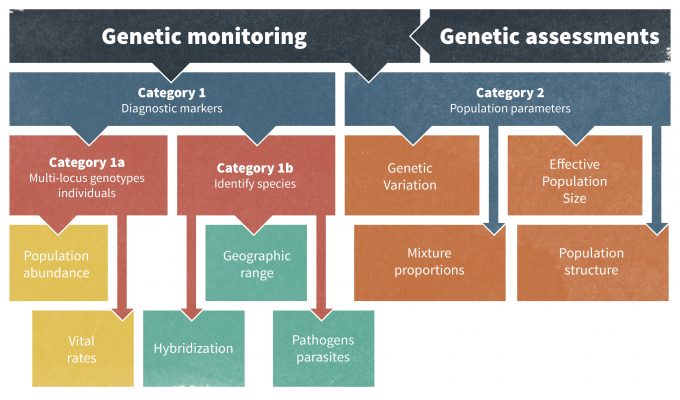
Un posible problema con la identificación de umbrales es que son el resultado de la negociación social y aunque pueden estar basados en la biología, también pueden representar simplemente un punto pactado, socialmente aceptable por directivos y actores. Los umbrales basados en la biología pueden representar densidad poblacional, probabilidad de ocurrencia, un cambio en la reproducción o supervivencia (o lambda), heterocigosidad genética u otros parámetros poblacionales, pero los umbrales son establecidos conjuntamente por biólogos y partes interesadas. El uso de marcadores genéticos para evaluar cambios en el tamaño efectivo de la población y otros aspectos de la ecología poblacional se ha vuelto cada vez más popular (Schwartz et al. 2007). Schwartz et al. (2007) describieron dos categorías de monitoreo usando marcadores genéticos: Categoría I que puede identificar individuos, poblaciones y especies; y Categoría II que monitorea los parámetros genéticos de la población permitiendo conocer los procesos demográficos y 'monitoreo retrospectivo' para comprender mejor cambios históricos (Figura 13.2). Sin embargo, los umbrales también pueden constituir un reflejo más de la tolerancia o los deseos de la sociedad por una especie en particular. Por ejemplo, el umbral para el número de pumas en una zona residencial de California puede ser el nivel que el público pueda tolerar más que lo que es más significativo en términos de la dinámica poblacional de la especie.
Tendencias de pronósticos
Con 5 años de datos, las tendencias pueden comenzar a emerger de los datos (Figura 13.3) que proporcionan información para guiar las acciones de gestión. En nuestro ejemplo hipotético, las tendencias en tierras públicas son bastante estables, mientras que las de las tierras privadas están disminuyendo. Si pronosticamos la tendencia de tierras privadas hacia el futuro podemos ver que en 2.5 años la x intercepción para la tendencia llegará a 0. El grado de precisión en la estimación de la intercepción x disminuye drásticamente a medida que los pronósticos se extienden más hacia el futuro, por lo que los intentos de pronóstico deben considerarse como una herramienta para guiar la toma de decisiones. No está claro si la intercepción x se alcanzará en 1 año, 2.5 años o 10 años, o en absoluto, pero la línea de tendencia sí genera preocupaciones sobre la viabilidad a largo plazo de la especie en tierras privadas y puede iniciar una respuesta más rápida que si la línea de tendencia tuviera una intercepción x de 15 años. Dunn (2002) utilizó un enfoque similar a este y clasificó a más de 200 especies de aves en categorías de alerta de conservación.
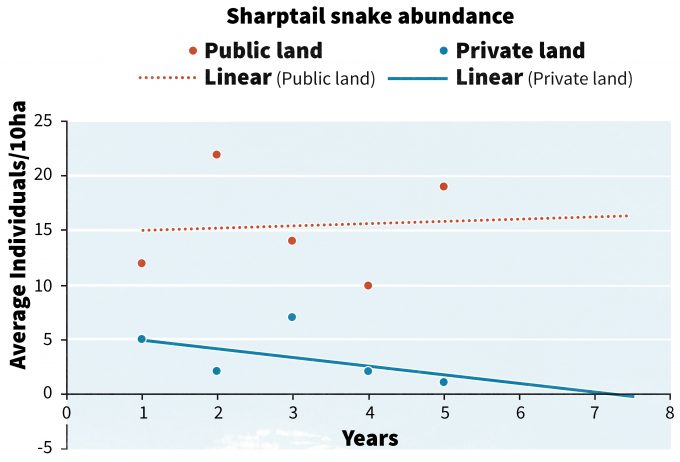
Pero estas son simplemente tendencias lineales y la variabilidad asociada a las tendencias, especialmente para especies raras, suele ser muy alta. En efecto, el poder asociado a detectar una tendencia significativa suele ser muy bajo con especies raras, por lo que las líneas de tendencia estadística deben interpretarse con cautela para evitar cometer un error de concluir que no existe tendencia cuando en realidad está en declive. Esto es especialmente problemático cuando las poblaciones ya han alcanzado niveles muy bajos y la probabilidad de detectar una disminución adicional es muy baja (Staples et al. 2005). En estos casos puede ser más útil emplear evaluaciones de riesgo basadas en análisis de viabilidad poblacional (PVA, Morris et al. 2002, Lande et al. 1993). Si los datos recopilados en el monitoreo pueden usarse para ayudar a parametrizar un modelo de PVA, entonces se pueden estimar al menos cambios relativos en la abundancia futura de la población o el tiempo de extinción (Dennis et al. 1991, Morris y Doak 2002). Staples et al. (2002) propusieron un enfoque de monitoreo poblacional viable en el que se utilizan predicciones de riesgo anuales como indicador de monitoreo. Staples et al. (2002) definieron 'riesgo' como “la probabilidad de que la abundancia poblacional disminuya por debajo de un umbral inferior dentro de un marco de tiempo dado”. Predecir que el riesgo aumentará con el tiempo podría constituir un punto desencadenante e impulsar acciones alternativas de gestión.
Predecir patrones sobre el espacio y el tiempo
Claramente a los gestores les gustaría saber en qué parte de un paisaje es probable que se produzcan las especies para que se puedan tomar acciones de manejo para aumentar, disminuir las poblaciones o al menos tener efectos mínimos sobre las especies deseadas. El monitoreo de la ocurrencia de organismos en un paisaje puede proporcionar información sobre la distribución espacial de individuos dentro de las poblaciones y puede proporcionar una mejor comprensión de la estructura de metapopulación y conectividad entre subpoblaciones. Si también se incluye información sobre reproducción y supervivencia en el esfuerzo de monitoreo, entonces también se puede obtener información adicional sobre el valor de las subpoblaciones como fuentes o sumideros. Y si esta información se recoge a lo largo del tiempo entonces la información sobre las probabilidades de que las subpoblaciones se extingan localmente en parches y la posterior recolonización también se puede entender a través de un monitoreo a largo plazo.
Si bien esta información de referencia sobre la distribución y adecuación de los organismos en un área de planeación es información valiosa para comprender los impactos del manejo de paisajes, temas como el uso del suelo y el cambio climático hacen que la información sea aún más valiosa. Ante tales cambios, el riesgo de pérdida de especies de un área, o incluso de extinción general, depende de la velocidad a la que una especie pueda adaptarse a las condiciones cambiantes. La información de monitoreo puede proporcionar evidencia para comprender mejor tanto las tasas de cambio en el ambiente biofísico como la aptitud asociada de los organismos. En los siguientes apartados utilizamos el monitoreo en el contexto del cambio climático como ejemplo para mostrar cómo los estresores ambientales pueden influir en cómo actúan los gerentes para intentar conservar la biodiversidad, pero también las dificultades de enfrentar cambios ecológicos tan integrales.
Si seguimos bombeando CO 2 a la atmósfera a las tasas actuales, entonces aproximadamente 20-30% de las especies vegetales y animales evaluadas por el IPCC (2007) probablemente estén en mayor riesgo de extinción a medida que la temperatura promedio global aumente en 1.5 a 2.5°C o más. Por lo tanto, comprender ciertos aspectos del ambiente y las respuestas de las especies a través del monitoreo es clave para brindar oportunidades para que las especies se adapten o se recuperen del cambio climático. Pero el cambio climático es probablemente uno de los estresores ambientales más difíciles de responder incluso con buenos datos de monitoreo porque es global; la oportunidad de comparaciones entre sitios afectados por el cambio climático y aquellos no afectados por el cambio es rara si existen. En efecto, no se nos suele dar la oportunidad de utilizar BACI o enfoques mensurativos comparativos al diseñar un plan de monitoreo que afecte a los estresores regionales o globales, por lo que debemos más bien confiar en las asociaciones a lo largo del tiempo. Para ser más específicos, el efecto puede inferirse de estos datos solo con cuidado ya que otros factores asociados al cambio pueden tener un mayor o menor efecto en cualquier tendencia observada. Sin embargo, hay una serie de factores potenciales que a menudo se evalúan al tratar de comprender los efectos de los cambios globales como el cambio climático en la pérdida de biodiversidad.
Cambios en el rango geográfico
Si los aumentos de temperatura promedio global superan 1.5 a 2.5°C, entonces se anticipan cambios importantes en la estructura y función de los ecosistemas, las interacciones ecológicas de las especies y los cambios en los rangos geográficos de las especies, con consecuencias predominantemente negativas
para la biodiversidad (IPCC 2007). Debido a que los rangos geográficos de las especies a menudo están dictados por condiciones climáticas (o por barreras topográficas) que influyen en las respuestas fisiológicas, los cambios en los rangos geográficos de las especies se predicen con frecuencia usando envolturas bioclimáticas (Pearson y Dawson 2003), y los cambios observados se utilizan como indicador de la respuesta de una especie y capacidad de adaptación al cambio climático. Pero las envolturas bioclimáticas están siendo examinadas y cuestionadas porque las interacciones bióticas, el cambio evolutivo y la capacidad de dispersión también influyen en la capacidad o incapacidad de una especie para responder a los cambios en su entorno (Pearson y Dawson 2003). Uno puede imaginar fácilmente cómo los impactos del cambio climático en las subpoblaciones podrían verse exacerbados por el cambio de uso de la tierra que conduzca a su aislamiento; de hecho, estas subpoblaciones se volverían más vulnerables a la extinción local por incapacidad para dispersarse, enfermedades infecciosas o competencia con especies invasoras como sus cambios de hábitat en respuesta al cambio climático.
Zuckerberg et al. (2009) utilizaron el New York State Breeding Bird Atlas encuestado en 1980-1985 y 2000—2005 para probar predicciones de que los cambios en la distribución de aves están relacionados con el cambio climático. Encontraron que 129 especies de aves mostraron un desplazamiento promedio hacia el norte en su latitud media de 3.6 km (Zuckerberg et al. 2009) y que los límites del rango sur de algunas especies de aves se movieron hacia el norte en 11.4 km. Claramente estos programas de monitoreo pueden proporcionar evidencia de asociaciones entre el cambio climático y los cambios en los rangos geográficos, sin embargo, no deben descartarse otros factores. La densidad de población humana ha cambiado a lo largo de ese tiempo al igual que los patrones de uso del suelo y ambos podrían haber tenido efectos similares en el rango geográfico de ciertas especies. Sin embargo, el hecho convincente es que todas las 129 especies que examinaron mostraron un desplazamiento hacia el norte en la distribución, por lo que en este caso, los datos sugieren que la influencia impulsora es algo más global y consistente en su impacto. Los esfuerzos similares en el uso de la información de monitoreo a lo largo del tiempo pueden dilucidar cambios para especies menos móviles como plantas, invertebrados y anfibios (Walther et al. 2002).
Inicio Gama Tamaños
La disponibilidad de recursos está relacionada con el tamaño del rango de hogares para muchas especies. Es muy probable que el cambio climático influya en la dispersión o concentración de los alimentos disponibles y cubra los recursos para muchas especies (McNab 1963). Por lo tanto, el monitoreo del tamaño del rango doméstico también constituye un método para evaluar los efectos ecológicos del cambio climático en algunas especies.
Documentar los tamaños de los rangos domésticos puede ser costoso y las estimaciones pueden sufrir de baja precisión por varias razones (Borger et al. 2006). Estimar el tamaño del efecto que podría detectarse (un análisis de potencia ayudaría a determinarlo, Zielinski y Stauffer 1996), puede permitir una mejor comprensión de los riesgos reales de perder especies.
Algunos efectos son evidentes en las latitudes más altas. A medida que el hielo marino se pierde y se desplaza en sus ubicaciones, los osos polares deben extender sus ataques de forrajeo a nuevas ubicaciones (Derocher et al. 2004). Si la energía que gastan en forrajeo excede la energía que obtienen al atrapar presas entonces morirán. Con osos polares y otras especies, la expansión del tamaño del área de distribución de los hogares puede ser un indicador de alerta temprana de disponibilidad de recursos disminuida o dispersa y una indicación de que la especie puede estar enfrentando una disminución inminente de la población. Por lo tanto, los cambios en los tamaños de los hogares pueden ser un aspecto importante del análisis de riesgos. Si los tamaños de los hogares se están expandiendo, entonces el riesgo de disminución de la población es mayor que si son estables a la contracción y los cambios en el tamaño del rango de esperanza pueden ser detectables antes de una disminución en la abundancia
Cambios fenológicos
Otra señal de alerta temprana de impactos inminentes del cambio climático en las poblaciones son los cambios en los patrones fenológicos de plantas y animales. De hecho, eventos como la llegada a sitios de reproducción o invernada a partir de migraciones, inicio de floración u otras actividades reproductivas, hojas o caída de hojas funcionan como indicadores porque tienden a verse influenciados al menos en parte por la temperatura (Parmesan 2007). Los estudios fenológicos se han realizado desde hace años (por ejemplo, Menzel 2000), pero no a escala global necesaria para monitorear el cambio climático global. Schwartz (1994) brindó una discusión sobre la detección de cambios a gran escala utilizando información fenológica. Comentó cómo los esfuerzos pasados para registrar patrones fenológicos a menudo se han realizado a pequeña escala, y luego sugirió que al integrar la recolección de datos terrestres con datos de detección remota, los patrones locales se pueden escalar apropiadamente a ecorregiones, continentes e idealmente el globo, permitiendo una mayor escala patrones a inferir. White et al. (2005) propusieron un marco global para monitorear las respuestas fenológicas al cambio climático utilizando datos de detección remota. Si el enfoque de White et al. (2005) se puede implementar de manera efectiva, entonces los mecanismos físicos responsables de los patrones observados pueden ser utilizados para evaluar la efectividad de los modelos a escala global en la predicción de cambios en eventos fenológicos (Schwartz 1994).
Estructura y composición del hábitat
Para algunos propósitos, simplemente comprender los cambios en la disponibilidad de hábitat para una especie puede ser suficiente para inferir cambios probables en la abundancia o distribución potencial de una especie. El diablo está en los detalles sin embargo. Para muchas especies, el conocimiento de la abundancia y la disposición espacial de elementos de hábitat de escala fina como árboles grandes, enganches, troncos o arbustos es importante. Pero reunir este conocimiento a gran escala puede plantear problemas; las imágenes satelitales no detectarán muchas de estas características. Sin embargo, LIDAR u otros datos de detección remota a menudo pueden proporcionar información a una escala lo suficientemente fina como para detectar componentes del hábitat (Hyde et al. 2006). LIDAR en particular puede proporcionar información sobre la complejidad vertical a escala fina de un bosque, incluyendo alturas de dosel y biomasa de dosel (Hyde et al. 2006). Para aquellas especies asociadas con capas vegetativas en bosques, los datos de detección remota pueden ser útiles. Para las especies asociadas con madera muerta u otros elementos de hábitat que no son detectables mediante técnicas remotas, entonces la combinación de datos de detección remota con datos de parcelas terrestres se convierte en el único enfoque lógico. Estos elementos de hábitat de escala fina se pueden imputar a píxeles de ubicaciones conocidas de parcelas terrestres utilizando técnicas de vecinos más cercanos (Ohmann y Gregory 2002).
Síntesis de Datos de Monitoreo
Los datos de monitoreo pueden integrarse con otra información sobre el terreno, el clima, las probabilidades de perturbación, el uso del suelo, la propiedad de la tierra y la infraestructura para pintar una imagen integrada generalizada del estado de un paisaje. Estos enfoques permiten a los gerentes monitorear no solo las piezas individuales del paisaje sino también el conjunto integrado a lo largo del tiempo. Por ejemplo, los cambios en la estructura y composición de rodales forestales en Oregón con y sin ciertas prácticas silvícolas pueden incorporarse a mapas de clases de edad forestal y tipos de hábitat (Spies et al. 2007). Estos pueden vincularse entonces a modelos de crecimiento y desarrollo forestal (muchos de los cuales se basan en parcelas de monitoreo continuo de inventarios forestales), y con probabilidades de transición asociadas a las decisiones de manejo de tierras, permitiendo proyecciones de posibles condiciones futuras con fines de planeación y para mejorar comprender las implicaciones de los posibles cambios en la política de uso del suelo (Spies et al. 2007). Otros enfoques no han utilizado explícitamente modelos de crecimiento de vegetación sino que han desarrollado escenarios de condiciones pasadas, condiciones actuales y condiciones futuras alternativas probables de paisajes (Baker et al. 2004). Es importante destacar que estos enfoques no solo utilizan información de monitoreo para parametrizar muchas de las proyecciones espaciales y temporales, sino también para mejorar nuestra comprensión de posibles condiciones futuras. En efecto, es la capacidad de utilizar datos para crear modelos que permitan proyecciones de condiciones hacia el futuro basadas en factores estresantes interactuantes como el cambio climático (IPCC 2007) y la planificación del uso del suelo (Kaiser et al. 1995). Estas proyecciones modelo no solo elevan el potencial para desarrollar escenarios “qué pasaría si” para comparar políticas alternativas, sino que también pueden identificar parámetros clave que deben ser monitoreados en el futuro para ayudar a las partes interesadas a comprender si los resultados de un cambio de política se están realizando como se proyecta. Hay tantas suposiciones interactuantes que entran en estas complejas proyecciones de paisaje que sin datos de monitoreo, las proyecciones son en el mejor de los casos una condición futura probable y en el peor de los casos un artefacto de una suposición incorrecta. Algunos profesionales también intentan integrar los datos de monitoreo ecológico con información económica, social e institucional para crear cuerpos de datos que funcionen como indicadores de sustentabilidad. Esto a menudo se ha hecho para sistemas agrícolas y para comunidades de países en desarrollo pero se está expandiendo para incluir otras regiones, como entornos urbanos altamente desarrollados (Olewiler 2006, Van Cauwenbergh et al. 2007). No todas estas iniciativas incluyen necesariamente el monitoreo de poblaciones o hábitat, pero muchas sí. Por ejemplo, para evaluar la sustentabilidad del uso de los recursos terrestres de las comunidades en ecosistemas tropicales, varios investigadores han integrado el monitoreo de vida silvestre y el mapeo de sitios de matanza de caza con datos sobre el uso de otros recursos terrestres, el acceso a nuevas tecnologías y los cambios locales usos del suelo (Koster 2008, Parry et al. 2009). En el marco de Evaluación de Sustentabilidad de la Agricultura y el Medio Ambiente (SAFE) para desarrollar un conjunto de variables que indiquen la sustentabilidad de los agroecosistemas, las variables que miden la retención de la biodiversidad y la “calidad funcional de los hábitats” se consideran un componente integral de la marco de monitoreo (Van Cauwenbergh et al. 2007). Si bien el monitoreo de la vida silvestre y el hábitat no se discute explícitamente dentro de los lineamientos marco, sería difícil hacer tales evaluaciones sin hacerlo. También es importante darse cuenta de que el concepto de “indicadores de sustentabilidad” y los intentos previos de derivarlos tiene su parte de críticos. Scerri y James (2009), por ejemplo, discuten cuántos practicantes reducen el complejo concepto de sustentabilidad y la generación de indicadores de sustentabilidad que probablemente sean específicos de contexto para una tarea muy técnica y cuantitativa.
Los modelos de PVA suelen comparar el riesgo estimado de que una especie o población se extinga entre varias alternativas de manejo. Los modelos de PVA son notoriamente hambrientos de datos que requieren estimaciones específicas de la edad o etapa de supervivencia, reproducción y movimientos con rangos de variabilidad asociados para cada estimación de parámetros (Beissenger y Westphal 1998, Reed et al. 2002). Al igual que con las proyecciones de modelos de paisaje, los aspectos de monitoreo de las proyecciones de PVA permiten no solo una evaluación del riesgo asociado a no lograr un resultado esperado sino también resaltar las debilidades en los supuestos del modelo. Los programas de monitoreo que informan la validez de los supuestos pueden brindar la oportunidad de desarrollar estructuras modelo más confiables y proyecciones resultantes. Decidir qué supuestos o parámetros monitorear con base en una estructura de modelo puede ser problemático, especialmente con modelos complejos grandes como los dos descritos anteriormente. La identificación de la variable a monitorear puede basarse en una evaluación subjetiva de la confiabilidad de los datos subyacentes o mediante análisis de sensibilidad más estructurados que identifican variables que tienen una influencia primordial en los resultados del modelo (McCarthy et al. 1995, Fieldings y Bell 1997). Muy a menudo los parámetros menos confiables en estos modelos son aquellos que son los más difíciles de medir. Esto puede crear un dilema para un gerente de programa que desarrolle un programa de monitoreo ya que estos datos pueden ser los más importantes para llevar a una disminución de la incertidumbre en predicciones futuras pero también pueden ser los más caros de adquirir. De ahí que sea necesario realizar una evaluación beneficio:costo con las partes interesadas para elaborar una lista prioritaria de variables.
A pesar de la capacidad de desarrollar estimaciones más confiables de variables clave a partir de datos de monitoreo, las proyecciones hacia el futuro siempre se enfrentan a la incapacidad de predecir eventos de umbral desconocidos que no se habrían previsto al principio. Por ejemplo, las invasiones de búhos excluidos en el hábitat del búho manchado no se consideraron seriamente una amenaza tanto como la pérdida de hábitat cuando se desarrollaron PVA tempranos para búhos manchados (Peterson y Robins 2003). E incluso cuando los modelos pueden considerar variables nuevas o confusas, las interrelaciones entre las variables pueden dar lugar a nuevos estados o procesos que no podrían preverse.
Los climas siempre han cambiado en esta tierra pero la tasa de cambio que probablemente se verá en el próximo siglo podría no tener precedentes. Es probable que los cambios en la estructura vegetativa de la comunidad y las relaciones interespecíficas cambien, pero su capacidad de adaptación a las cambiantes condiciones climáticas está en duda. Williams y Jackson (2007) proporcionaron una visión general de las comunidades de plantas no analógicas asociadas con climas históricos “novedosos” y futuros climas novedosos que probablemente sean más cálidos que cualquier otro presente. Los modelos ecológicos como los modelos de dinámica forestal y los modelos PVA se parametrizan al menos parcialmente a partir de datos recopilados relativamente recientemente, por lo que es posible que no predigan con precisión las respuestas a climas novedosos (Williams y Jackson 2007). La incertidumbre generada por el desarrollo potencial de condiciones no analógicas debe considerarse explícitamente durante los análisis de riesgo.
Análisis de Riesgos
Se han desarrollado formalmente análisis de riesgos con respecto a los efectos directos e indirectos de los contaminantes en las especies de vida silvestre. La Agencia de Protección Ambiental define la Evaluación de Riesgos Ecológicos (EEI) como, “una evaluación de los posibles efectos adversos que las actividades humanas tienen sobre los organismos vivos que conforman los ecosistemas. El proceso de evaluación de riesgos proporciona una forma de desarrollar, organizar y presentar información científica para que sea relevante para las decisiones ambientales. Cuando se realiza para un lugar en particular, como una cuenca hidrográfica, el proceso de EEI puede usarse para identificar recursos vulnerables y valorados, priorizar la actividad de recolección de datos y vincular las actividades humanas con sus posibles efectos. Los resultados de ERA proporcionan una base para comparar diferentes opciones de manejo, permitiendo a los tomadores de decisiones y al público tomar decisiones mejor informadas sobre el manejo de los recursos ecológicos” (http://epa.gov/superfund/programs/nrd/era.htm). Los pasos utilizados por la EPA se describen en la Figura 13.4, y podrían adaptarse para su uso en otras situaciones donde los riesgos de otros estresores o perturbaciones ambientales pueden ser de importancia clave para los gerentes (por ejemplo, incendios, uso del suelo, inundaciones, etc.). Por ejemplo, Hull y Swanson (2006) proporcionaron un proceso paso a paso para evaluar el riesgo para las especies de vida silvestre por la exposición a contaminantes. Se han propuesto enfoques similares para evaluar el riesgo de pérdida de biodiversidad. Kerns y Ager (2007) describieron la evaluación de riesgos como un procedimiento para evaluar amenazas y comprender la incertidumbre “... proporcionando: (1) una estimación de la probabilidad y gravedad de la pérdida o ganancia de especies, poblaciones o hábitats, (2) una mejor comprensión de las posibles compensaciones asociadas con las actividades de manejo, y 3) integración socioeconómica tangible.” Propusieron una evaluación de riesgos cuantitativa y probabilística para proporcionar un puente entre la planificación y la política que incluya la participación de las partes interesadas (Kerns y Ager 2007). Dichos enfoques formales son necesarios dentro de los procesos de planeación ecológica para que tanto los gerentes como las partes interesadas entiendan la incertidumbre y los costos asociados con los riesgos de no lograr los resultados previstos.
Toma de Decisiones
Desde un punto de vista lógico, las decisiones deben tomarse mediante una secuencia de pasos: caracterizar el problema o pregunta, identificar una gama completa de alternativas y determinar
criterios para seleccionar una, recopilar información sobre cada opción y calificarla según los criterios, luego tomar la decisión final con base en la calificación (Lach y Duncan 2007). Pero Klein (2001) encontró que solo el 5% de todas las decisiones se toman utilizando un enfoque tan lógico. Los individuos suelen tomar sus decisiones utilizando la intuición y simulaciones mentales (relacionando rápidamente el resultado de una decisión con alguna experiencia) (Lach y Duncan 2007). Los grupos pueden tomar decisiones de manera diferente y los grupos son más capaces de tomar mejores decisiones sobre problemas complejos que los individuos (Lach y Duncan 2007). Las personas con diferentes visiones del mundo estructuran el mundo que les rodea de diferentes maneras y al hacerlo aportan una perspectiva diferente a una decisión grupal. Asegurar que una variedad de visiones del mundo estén representadas en un grupo puede ser particularmente útil cuando se trata de llegar a una decisión equilibrada sobre un tema complejo, aunque las discusiones necesarias para llegar a esa decisión necesariamente pueden llegar a ser prolongadas.
Resumen
Se invierte considerable tiempo y dinero en muchos programas de monitoreo por lo que no sólo el diseño de estos programas debe ser científicamente y estadísticamente riguroso, sino que debe tener claro para los directivos y partes interesadas cómo se utilizará la información para tomar decisiones. Durante la fase de diseño, se deben identificar los puntos de activación o umbrales para asegurar que los gerentes sepan cuándo deben considerarse los cambios en los enfoques de gestión. En muchas circunstancias es fácil para los directivos simplemente esperar más información sin tomar una acción, sin darse cuenta de que la espera pone mayor riesgo en el logro de los resultados deseados. El uso de datos de monitoreo como base para pronosticar tendencias en el espacio y el tiempo puede permitir a los gerentes comprender las implicaciones de esperar demasiado tiempo antes de tomar medidas correctivas. Factores como cambios en las características climáticas, fenología, rangos geográficos y tamaños de rango de hogares de algunas especies pueden ser particularmente informativos ante los cambios climáticos globales para los que la única condición de referencia es el pasado. El uso de la información de monitoreo como medio de parametrización de modelos de paisaje o cambio climático permite proyecciones sobre el espacio y tiempos de condiciones más complejas. Dichos enfoques integradores permiten además comparaciones entre estrategias o políticas alternativas de manejo y pueden ser un componente importante de un análisis de riesgos, un enfoque formalizado para identificar incertidumbres y evaluar los efectos directos e indirectos del estrés en organismos y ecosistemas. Los resultados de monitoreo, modelado y análisis de riesgos se utilizan luego para tomar decisiones por individuos o por grupos. Aunque normalmente asumimos que las decisiones se toman de manera lógica, muchas decisiones se toman basadas en la intuición o como resultado de discusiones grupales entre personas con diversas visiones del mundo.
Referencias
Baker, J.P., D.W. Hulse, S.V. Gregory, D. White, J. Van Sickle, P.A. Berger, D. Dole, y N.H. Schumaker. 2004. Futuros alternativos para la cuenca del río Willamette. Aplicaciones Ecológicas 14:313 —324.
Beissinger, S.R. y M.I.Westphal. 1998. Sobre el uso de modelos demográficos de viabilidad poblacional en el manejo de especies amenazadas. Revista de Manejo de Vida Silvestre 62:821-841.
Block, W.M., A.B. Franklin, J.P. Ward Jr., J.L. Ganey, y G.C. White. 2001. Diseño e implementación de estudios de monitoreo para dilucidar el éxito de la restauración ecológica en la vida silvestre. Ecología de Restauración 9:293 —303.
Borger, L., N. Franconi, F. Ferretti, F. Meschi, G. De Michele, A. Gantz, A. Manica, S. Lovari, y T. Coulson. 2006. Efectos del régimen de muestreo sobre la media y varianza de las estimaciones de tamaño del rango de hogares. Revista de Ecología Animal 75:1393 —1405.
Dennis, B., P.L. Munholland, y J.M. Scott. 1991. Estimación de parámetros de crecimiento y extinción de especies amenazadas. Monografías Ecológicas 61:115 —143.
Derocher, A., Lunn, N.J., Stirling, I. 2004. Osos polares en un clima cálido. Biología Comparada Integrativa 44:163 —176.
Dunn, E.H., 2002. Utilizar la disminución de las poblaciones de aves para identificar las necesidades de acción de conservación. Biología de la Conservación 16:1632 —1637.
Fieldings, A.H., y J.F. Bell. 1997. Revisión de métodos para la evaluación de errores de predicción en presencia de conservación: modelos de ausencia. Conservación Ambiental 24:38—49.
Hull R.N., y S. Swanson. 2006. Análisis secuencial de líneas de evidencia: un enfoque avanzado de ponderación de evidencia para la evaluación de riesgos ecológicos. Evaluación y Gestión Ambiental Integrada 2:302 —311.
Hyde, P., R. Dubayah, W. Walker, J.B. Blair, M. Hofton y C. Hunsaker. 2006. Mapeo de la estructura forestal para el análisis de hábitat de vida silvestre usando sinergia multisensor (LiDAR, SAR/InSAR, ETM+, Quickbird). Teledetección del Medio Ambiente 102:63-73
IPCC. 2007. Cambio climático 2007: informe de síntesis. Contribución de los grupos de trabajo I, II y III al cuarto informe de evaluación del Panel Intergubernamental de Cambio Climático. IPCC, Ginebra, Suiza, 104 pp.
Kaiser, E., D. Godschalk, y F.S. Chapin. 1995. Planeación urbana del uso del suelo. Prensa de la Universidad de Illinois, Cuarta Edición. Urbana, IL.
Kerns, B.K. y A. Ager. 2007. Evaluación de riesgos para la planificación de conservación de la biodiversidad en bosques del noroeste del Pacífico Ecología y Manejo Forestal 246:38-44.
Klein, G. 2001. Comprender y apoyar la toma de decisiones: Una entrevista con Gary Klein. Gestión de Sistemas de Conocimiento de la Información 2 (4) :291—296.
Koster, J. 2008. El impacto de la caza con perros en las cosechas de vida silvestre en la Reserva Bosawas, Nicaragua. Conservación Ambiental 35 (3) :221-220.
Lach, D. y S. Duncan. 2007. ¿Cómo tomamos decisiones? Capítulo 2, páginas 12-20 en Johnson, K.N., S. Gordon, S. Duncan, D. Lach, B. McComb y K. Reynolds. Conservación de criaturas del bosque: una guía para la toma de decisiones y modelos de decisión para la biodiversidad forestal. Comisión Nacional de Ciencia para la Silvicultura Sustentable Informe final. NCSSF, Washington, DC.
Lande, R. 1993. Riesgos de extinción poblacional por estocásticas demográficas y ambientales y catástrofes aleatorias. El naturalista americano 142: 911—927.
McCarthy, M.A., M.A. Burgman, y S. Ferson. 1995. Análisis de sensibilidad para modelos de viabilidad poblacional. Conservación Biológica 73:93-100.
McNab, B.K. 1963. La bioenergética y la determinación del tamaño del rango del hogar. El naturalista americano 97:133-141.
Menzel A. 2000. Tendencias en las fases fenológicas en Europa entre 1951 y 1996. Revista Internacional de Biometeorología 44:76 —81.
Morris, W.F., P.L. Bloch, B.R. Hudgens, L.C. Moyle, y J.R. Stinchcombe. 2002. Análisis de viabilidad poblacional en planes de recuperación de especies amenazadas: uso pasado y mejoras futuras. Aplicaciones Ecológicas 12: 708—712.
Morris, W.F., y D.F. Doak. 2002. Biología cuantitativa de la conservación: teoría y práctica del análisis de viabilidad poblacional. Asociados Sinauer, Sunderland, MA.
Ohmann, J.L. y Gregory, M.J., 2002. Mapeo predictivo de composición y estructura forestal con análisis de gradiente directo e imputación de vecinos más cercanos en la costa de Oregón, Estados Unidos. Revista Canadiense de Investigación Forestal 32:725-741.
Olewiler, N. 2006. Sustentabilidad ambiental para áreas urbanas: el papel de los indicadores de capital natural. Ciudades 23 (3) :184-195.
Parmesano, C. 2007. Influencias de especies, latitudes y metodologías en estimaciones de respuestas fenológicas al calentamiento global. Biología del Cambio Global 13:1860 —1872.
Parry, L., J. Barlow, y C.A. Peres. 2009. Asignación del esfuerzo de caza por parte de pequeños productores amazónicos: Implicaciones para la conservación de la vida silvestre en paisajes de uso mixto. Conservación Biológica. 142:1777 —1786
Pearson, R.G., y T.P. Dawson. 2003. Predecir los impactos del cambio climático en la distribución de las especies: ¿son útiles los modelos de envoltura bioclimática? Ecología Global y Biogeografía 12:361-371.
Peterson, A.T. y C.R. Robins. 2003. Usando modelos de nicho ecológico para predecir invasiones de búhos bloqueados con implicaciones para la conservación del búho manchado. Biología de la Conservación 17:1161 —1165.
Reed, J.M., L.S., Mills, J.B. Dunning, E.S. Menges, K.S.McKelvey, R. Frye, S.R. Beissinger, M.C. Anstett, y P. Miller. 2002. Temas emergentes en el análisis de viabilidad poblacional. Biología de la Conservación 16:7—19.
Scerri, A. y P. James. 2009. Comunidades de ciudadanos e 'indicadores' de sustentabilidad. Revista de Desarrollo Comunitario. doi:10.1093/cdj/bsp013
Schwartz, M.D. 1994. Monitoreo del cambio global con fenología: El caso de la ola verde primaveral. Revista Internacional de Biometeorolología 38:18 —22.
Schwartz M.K., G. Luikart, y R.S. Waples. 2007. El monitoreo genético como una herramienta prometedora para la conservación y manejo. Tendencias en Ecología y Evolución 22:25 —33.
Spies, T.A., K.N. Johnson, K.M. Burnett, J.L. Ohmann, B.C. McComb, G.H. Reeves, P. Bettinger, J.D. Kline, y B. Garber-Yonts. 2007. Efectos ecológicos y socioeconómicos acumulados de las políticas forestales en la costa de Oregón. Aplicaciones Ecológicas 17:5 —17.
Grapas D.F., M.L. Cono, y B.B. Shepard. 2005. Monitoreo poblacional viable basado en riesgos, Conservación. Biología 19:1908 —1916.
Van Cauwenbergh, N., K. Biala, C. Bielder, V. Brouckert, L. Franchois, V. García Caridad, M. Hermy, E. Mthij, B. Muys, J. Rejinders, X. Sauvenier, J. Valckx, M. Vancloster, B. Van der Veken, E. Wauters, y A. Peeters. 2007. Safe—un marco jerárquico para evaluar la sustentabilidad de los sistemas agrícolas. Agricultura, Ecosistemas y Medio Ambiente 120:229 —242.
Walther, G.-R., E. Post, P. Convey, A. Menzel, C. parmesano, T.J C. Beebee, J.-M. Fromentin, O. Hoegh-Guldberg, y F. Bairlein, 2002. Respuestas ecológicas al cambio climático reciente. Naturaleza 416:389 —395.
White, M.A., F. Hoffman, W.W. Hargrove, y R.R. Nemani. 2005. Un marco global para el monitoreo de las respuestas fonológicas al cambio climático. Letras de Investigación Geofísica 32:L04705.
Williams, J. W., y S. T. Jackson. 2007. Nuevos climas, comunidades no analógicas y sorpresas ecológicas. Fronteras en Ecología y Medio Ambiente 5:475 —482.
Zielinski, W.J., y H.B. Stauffer. 1996. Monitoreo de poblaciones de Martes en California: diseño de encuestas y análisis de potencia. Aplicaciones Ecológicas 6:1254 —1267
Zuckerberg, B., A.M. Woods y W.F. Porter. 2009. Cambios en la distribución de aves reproductoras en el estado de Nueva York. Biología del Cambio Global 15:1-18