
3.3: Inactivación de X
- Page ID
- 57536
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Introducción
Tener cromosomas adicionales o faltantes no suele ser algo bueno. De hecho, para la mayoría de los cromosomas, tener una copia extra o faltante es letal para los humanos (haciendo que un embrión muera temprano en el desarrollo).
Sin embargo, las hembras humanas tienen dos cromosomas X (XX), mientras que los machos humanos tienen solo uno (XY). ¿Por qué no causa problemas para los hombres tener solo una copia del cromosoma X, mientras que las mujeres tienen dos?
X-inactivación
Resulta que el nivel de actividad génica producida por un solo cromosoma X es la “dosis” normal para un ser humano. Los hombres tienen esta dosis porque, bueno, ¡solo tienen un cromosoma X! Las mujeres tienen la misma dosis por una razón diferente: cerraron uno de sus dos cromosomas X en un proceso llamado inactivación de X.
En la inactivación X, se compacta un cromosoma X (o, como me gustaba decir a mi bioprofesor de introducción, “arrugado en una bola”), para hacer una estructura pequeña y densa llamada cuerpo Barr. La mayoría de los genes en el cuerpo Barr son inactivos, es decir, que no se transcriben. El proceso de inactivación de X fue descubierto por la genetista británica Mary F. Lyon y a veces se le llama lionización en su honor 1.

Una mujer tiene dos cromosomas X, uno de cada padre. ¿Cuál va a inactivar? La inactivación de X es un proceso aleatorio que ocurre por separado en células individuales durante el desarrollo embrionario. Una célula podría cerrar la X paterna, mientras que su vecino de al lado podría cerrar la X materna en su lugar. Todas las células que descienden de cada una de estas células originales mantendrán el mismo patrón de inactivación de X.
Nota interesante: si fueras canguro, ¡lo que acabo de decir no sería verdad! En los canguros y otros marsupiales, siempre es el cromosoma X paterno el que sufre la inactivación de X 2.
Ejemplo de inactivación de X: Calico cat
Un ejemplo clásico de inactivación de X se observa en gatos. Si una gata hembra es heterocigota para los alelos negros y bronceados de un gen de color de pelaje que se encuentra en la X, inactivará sus dos X (y así, los dos alelos del gen del color del pelaje) al azar en diferentes células durante el desarrollo.
El resultado es un patrón de abrigo de carey, conformado por parches alternados de pelaje negro y bronceado. Los parches negros provienen de grupos de células en los que está activa la X con el alelo negro, mientras que los parches bronceados provienen de células en las que está activa la X con el alelo bronceado.
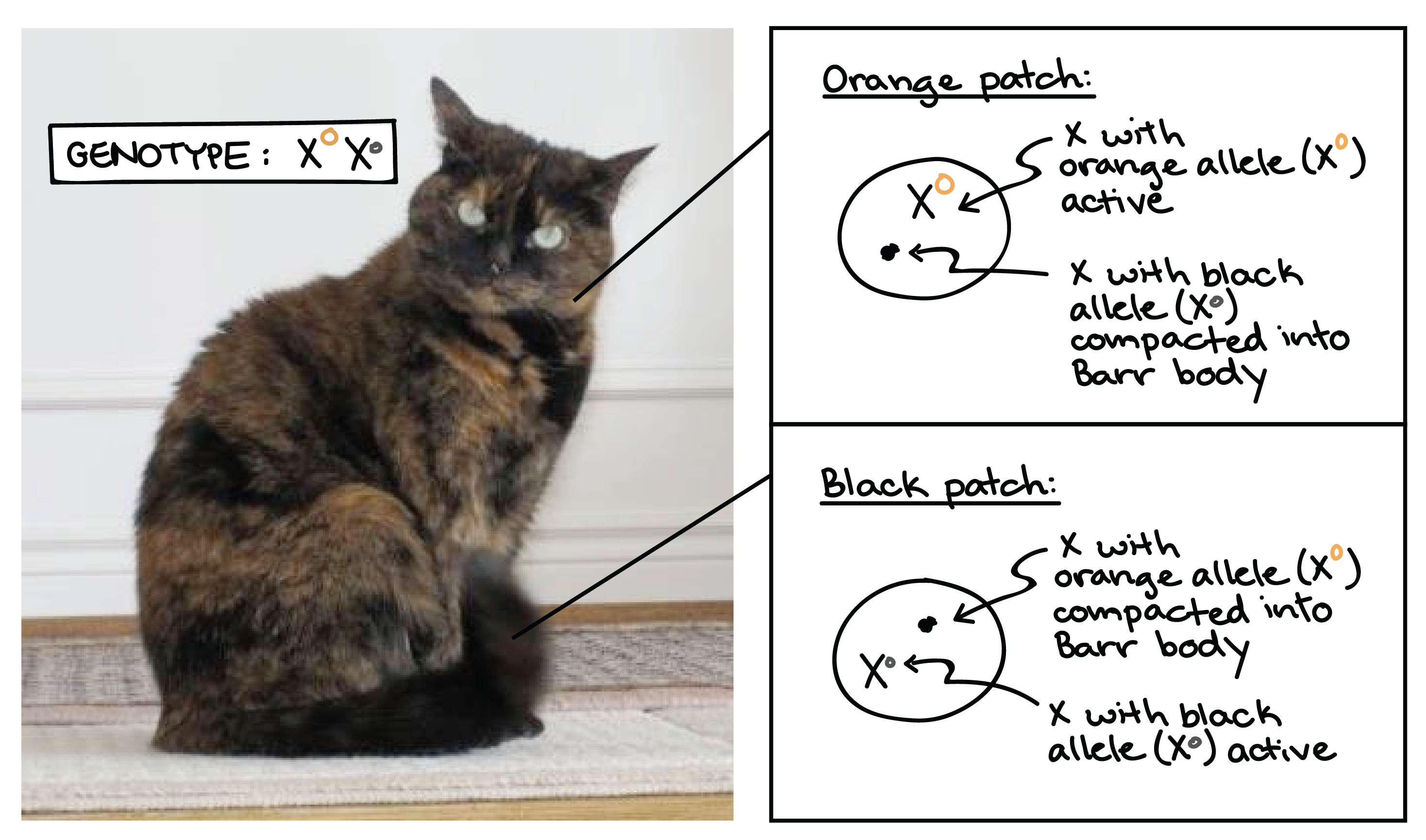
Aunque rara vez es tan fácil de ver como en el caso del gato carey, las hembras humanas también son “mosaico” para cualquier gen que esté presente en diferentes alelos en sus dos cromosomas X.
- [Si eso es cierto, ¿por qué las mujeres portadoras no muestran trastornos ligados al cromosoma X?]
-
En este punto, tal vez se esté preguntando por qué las mujeres heterocigóticas para un alelo recesivo ligado al X no muestran la condición asociada. Después de todo, aproximadamente la mitad de sus células inactivarán el alelo normal, dejando solo activo el alelo causante de trastorno.
En muchos casos, parece que tener el 50% de las células con una copia normal de un gen es suficiente para producir un fenotipo normal, o cercano a lo normal.
Un buen ejemplo de esto proviene de la condición ligada al X de deficiencia de visión de color rojo-verde (daltonismo). Las mujeres que son portadoras de alelos con deficiencia de visión de color rojo-verde tienen una mezcla de células fotorreceptoras activas y no trabajadoras en sus ojos. Sin embargo, las células de trabajo les permiten percibir el rojo y el verde lo suficientemente bien como para funcionar normalmente en la vida diaria. En pruebas más sensibles en un entorno de laboratorio, estas mujeres realmente muestran sutiles deficiencias de visión de color en relación con las mujeres que no son portadoras 3.
Aneuploidías de cromosomas sexuales
Cuando un organismo tiene una copia extra o faltante de un cromosoma, se dice que es aneuploide. Las aneuploidías que involucran autosomas (cromosomas no sexuales), especialmente las grandes, suelen ser tan dañinas para el desarrollo que un embrión aneuploide no puede sobrevivir hasta nacer.
Las aneuploidías de los cromosomas X, sin embargo, tienden a ser mucho menos dañinas, a pesar de que el X es un cromosoma grande. Esto es principalmente gracias a la inactivación de X. Si bien el propósito del sistema de inactivación X es apagar la segunda X de una hembra XX, también puede hacer un trabajo bastante bueno al apagar más cromosomas X si están presentes.
Los ejemplos de aneuploidías del cromosoma X incluyen:
- Síndrome Triple X, en el que una mujer tiene un genotipo XXX, el cual se presenta en aproximadamente 1 de cada 1,000 recién nacidas femeninas 4. Las mujeres con un genotipo XXX tienen características sexuales femeninas y son fértiles (capaces de tener hijos). En algunos casos, el síndrome triple X puede estar asociado con dificultades de aprendizaje, desarrollo tardío de habilidades motoras en infantes y problemas con el tono muscular 4.
- Síndrome de Klinefelter, en el que los machos tienen un cromosoma X extra, lo que lleva a un genotipo de XXY. (En casos más raros, el síndrome de Klinefelter puede involucrar varias Xs adicionales, lo que lleva a un genotipo XXXY o XXXXY). Los hombres afectados pueden ser infértiles o desarrollar vello corporal y facial menos denso que otros hombres. Se cree que el síndrome de Klinefelter afecta a 1 de cada 500 a 1,000 recién nacidos varones 5.
Al igual que las hembras, los machos XXY con síndrome de Klinefelter convertirán una X en un cuerpo de Barr en cada célula. Las hembras Triple X (así como los machos Klinefelter con más de dos cromosomas X) neutralizan sus Xs extra formando cuerpos Barr adicionales. Por ejemplo, habría dos cuerpos de Barr en una celda de una hembra XXX o un macho XXXY.

En el síndrome de Turner, una mujer carece de parte o la totalidad de uno de sus cromosomas X (dejándola con una sola X funcional). Las personas con este trastorno se desarrollan como mujeres, pero a menudo tienen baja estatura y pueden presentar síntomas como infertilidad y dificultades de aprendizaje. Se cree que el síndrome de Turner ocurre en aproximadamente 1 de cada 2,500 nacimientos femeninos 6. Tiene efectos relativamente leves porque los humanos normalmente tienen solo una X activa en las células de su cuerpo de todos modos.
Verifica tu comprensión
Consulta\(\PageIndex{1}\)
- [Pista]
-
Los gatos carey suelen tener un patrón de manchas mixtas de negro y naranja en sus pelajes como resultado de la inactivación de X. En gatos, un gen portado en el cromosoma X controla el color del pelaje, y las hembras heterocigóticas con un alelo naranja (\(\text{X}^O\)) y un alelo negro (\(\text{X}^o\)) inactivarán aleatoriamente uno de los dos cromosomas X en cada célula durante el desarrollo embrionario. Todas las células que descienden de cada una de estas células tempranas conservarán el mismo estado de inactivación, dando lugar a parches de células con la misma X activa y, por lo tanto, el mismo gen activo para la pigmentación de la capa.
Normalmente, los gatos machos no muestran un patrón de pelaje de carey porque solo tienen un cromosoma X, y por lo tanto no llevan a cabo la inactivación de X. En cambio, simplemente muestran el color sólido del pelaje correspondiente al alelo del gen del color del pelaje que se encuentra en su única X (con\(\text{X}^O\) dar naranja y\(\text{X}^o\) dar negro). Sin embargo, el sistema de inactivación de X todavía está presente en los machos, aunque normalmente no lo usan, y puede entrar en juego si un macho recibe un número inusualmente alto de cromosomas X.
Es posible obtener un patrón de carey en un gato macho si el gato recibe dos cromosomas X así como una Y, y si sus dos Xs llevan diferentes alelos del gen del color del pelaje (genotipo\(\text{X}^O\text{X}^o\text{Y}\)). Un genotipo XXY puede surgir por no disyunción, el fracaso de los cromosomas sexuales para separarse correctamente entre sí durante la meiosis. Un ejemplo de cómo la no disyunción podría conducir a un genotipo XXY y un fenotipo de carey se muestra en el cuadrado de Punnett debajo.
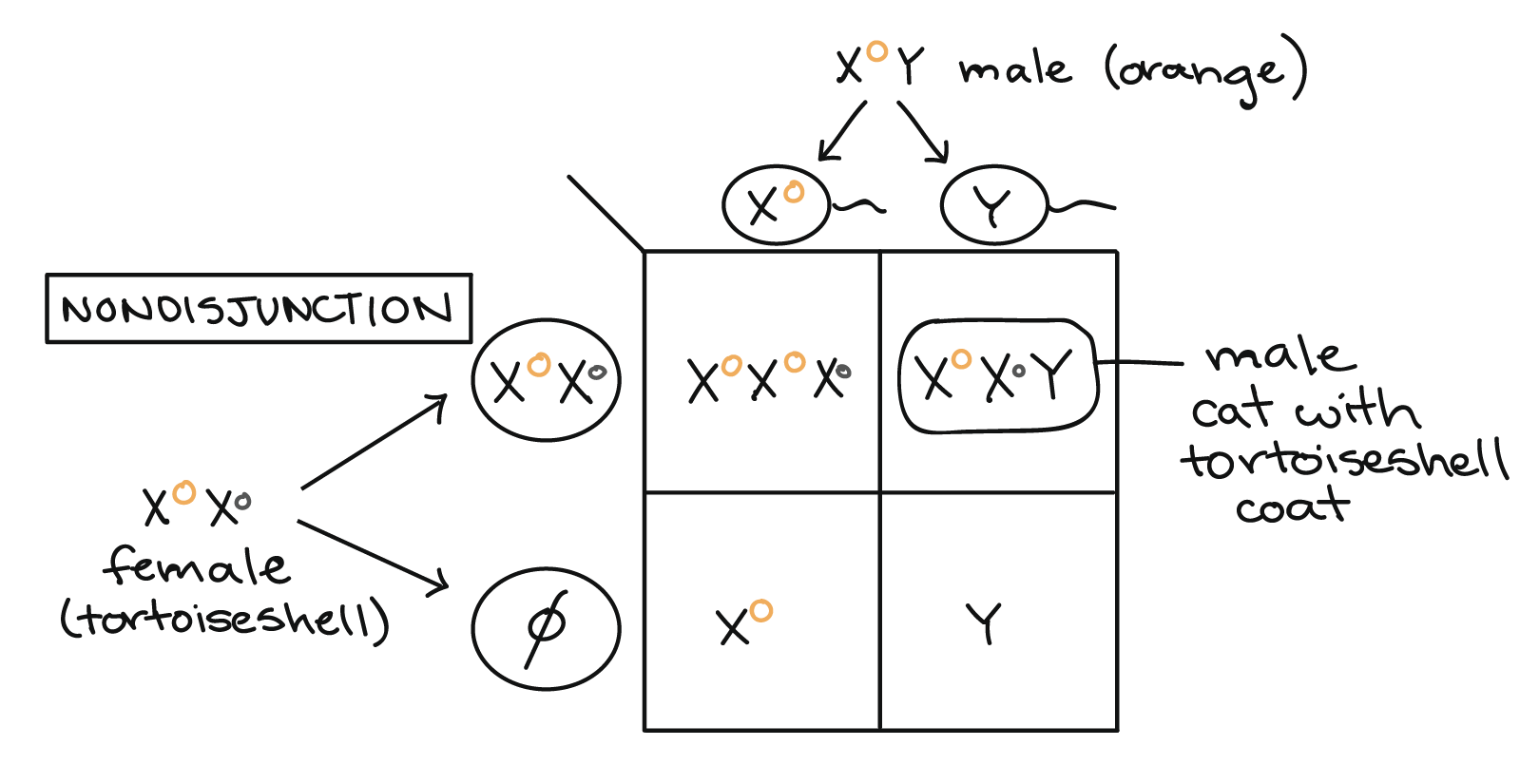
En gatos (como en humanos), el desarrollo masculino es especificado por el gen SRY en el cromosoma Y, por lo que un individuo XXY se desarrollará como macho. Sin embargo, solo una X puede estar activa en cada célula, por lo que uno de los dos cromosomas X se inactivará aleatoriamente en cada una de las células del macho XXY durante el desarrollo embrionario temprano. Esto da como resultado un patrón de capa de carey irregular como el de una gata hembra heterocigótica.
Las tortugas macho con genotipo XXY son externamente masculinas, pero suelen ser estériles (incapaces de tener descendencia), con una versión felina del trastorno conocido como síndrome de Klinefelter en humanos. Sin embargo, no todos los gatitos machos de carey tienen un número anormal de cromosomas X. En cambio, algunas son quimeras, o mosaicos genéticos, que resultan de la fusión de dos embriones en uno durante el desarrollo temprano. Si los dos embriones que se fusionan tienen diferentes alelos del gen del color del pelaje, el embrión quimérico tendrá una mezcla de células con diferentes genotipos, y el gato maduro tendrá pelaje irregular similar al de una carey hembra.
Colaboradores y Atribuciones
- [Atribución y referencias]
-
Atribución:
Este artículo es un derivado modificado de "Bases cromosómicas de los trastornos hereditarios”, por OpenStax College, Biology, CC BY 4.0. Descarga gratis el artículo original en http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.53.
Este artículo está bajo una licencia CC BY-NC-SA 4.0.
Obras citadas:
-
Mary F. Lyon. (2015, 22 de noviembre). Recuperado el 12 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Mary_F._Lyon.
-
Kimball, J. W. (2015, 23 de diciembre). Cromosomas sexuales. En las páginas de biología de Kimball. Recuperado de http://www.biology-pages.info/S/SexChromosomes.html.
-
Chen, Shuai. (2010, 4 de marzo). Dado que solo uno de los cromosomas X de una mujer funciona en una célula, ¿por qué no hay más mujeres daltónicas? [respuesta]. En Stanford en el Tech: Entendiendo la genética. Recuperado de http://genetics.thetech.org/ask/ask349.
-
Síndrome Triple X. (2014). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/condition/triple-x-syndrome.
-
Síndrome de Klinefelter. (2013). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/condition/klinefelter-syndrome.
-
Síndrome de Turner. (2012). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/condition/turner-syndrome.
Referencias adicionales:
Bergmann, D. C. (2011). Mutaciones, aneuploidía y determinación del sexo. En Genética notas de conferencia (pp. 36-44). Biosci41, Universidad de Stanford.
Chen, Shuai. (2010, 4 de marzo). Dado que solo uno de los cromosomas X de una mujer funciona en una célula, ¿por qué no hay más mujeres daltónicas? [respuesta]. En Stanford en el Tech: Entendiendo la genética. Recuperado de http://genetics.thetech.org/ask/ask349.
Deeb, S. S. y Motulsky, A. G. (2015, 5 de febrero). Defectos de visión de color rojo-verde. En GenereViews. Recuperado de http://www.ncbi.nlm.nih.gov/books/NBK1301/.
Factores genéticos y hormonas que determinan el género. (2007, 27 de junio). En Embriología Humana: Organogénesis. Recuperado de http://www.embryology.ch/anglais/ugenital/molec02.html.
Kimball, J. W. (2015, 23 de diciembre). Cromosomas sexuales. En las páginas de biología de Kimball. Recuperado de http://www.biology-pages.info/S/SexChromosomes.html.
Síndrome de Klinefelter. (2013). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/condition/klinefelter-syndrome.
Krempels, D. M. (n.d.). La genética de los gatos calicó. Recuperado de www.bio.miami.edu/dana/dox/calico.html.
Mary F. Lyon. (2015, 22 de noviembre). Recuperado el 12 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Mary_F._Lyon.
Colegio OpenStax, Biología. (2015, 13 de mayo). Teoría cromosómica y vinculación genética. En OpenStax CNX. Recuperado a partir de http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@9.85:64/Chromosomal-Theory-and-Genetic
O'Neil, Dennis. (2013). Anomalías del cromosoma sexual En Anomalías cromosómicas humanas. Recuperado de http://anthro.palomar.edu/abnormal/abnormal_5.htm.
OPN1LW. (2015). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/gene/OPN1LW.
Purves, W. K., Sadava, D. E., Orians, G. H., y Heller, H.C. (2003). Determinación del sexo y herencia ligada al sexo. En la vida: La ciencia de la biología (7ª ed., pp. 125-144). Sunderland, MA: Asociados Sinauer.
Reece, J. B., Urry, L. A., Caín, M. L., Wasserman, S. A., Minorsky, P. V., y Jackson, R. B. (2011). Las alteraciones del número o estructura cromosómica provocan algunos trastornos genéticos. En biología Campbell (10a ed., pp. 304-307). San Francisco, CA: Pearson.
Reece, J. B., Urry, L. A., Caín, M. L., Wasserman, S. A., Minorsky, P. V., y Jackson, R. B. (2011). Los genes ligados al sexo exhiben patrones únicos de herencia. En biología Campbell (10a ed., pp. 205-209). San Francisco, CA: Pearson.
Factor determinante de testículos. (2015, 10 de noviembre). Recuperado el 11 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/Testis_determining_factor.
Síndrome Triple X. (2014). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/condition/triple-x-syndrome.
Síndrome de Turner. (2012). En Genética hogar de referencia. Recuperado de http://ghr.nlm.nih.gov/condition/turner-syndrome.
Sistema de determinación del sexo XY. (2015, 19 de noviembre). Recuperado el 11 de diciembre del 2015 de Wikipedia: https://en.wikipedia.org/wiki/XY_sex-determination_system.
-