
3.4: Revisión de vinculación sexual




- Última actualización
- 29 oct 2022
- Guardar como PDF
( \newcommand{\kernel}{\mathrm{null}\,}\)
Términos clave
| Término | Significado |
|---|---|
| Cromosoma sexual | Uno de los dos cromosomas que determina el sexo biológico de un organismo |
| Autosome | Cromosoma que no es un cromosoma sexual |
| Gen ligado al sexo | Gen que se encuentra en uno de los dos cromosomas sexuales |
| Transportista | Individuo heterocigótico que heredó un alelo recesivo por un trastorno genético pero que no presenta síntomas de ese trastorno |
| Cuerpo Barr | Una región condensada en el núcleo de una célula, que consiste en un cromosoma X inactivado |
| Aneuploidía | Condición de tener demasiados o muy pocos cromosomas |
Vinculación sexual
En humanos, el sexo biológico está determinado por un par de cromosomas sexuales: XX en hembras y XY en machos. Los otros 44 cromosomas son autosomas.
Los genes en el cromosoma X o Y son rasgos vinculados al sexo. Los genes que se encuentran en el cromosoma X se pueden encontrar tanto en machos como en hembras, mientras que los genes que se encuentran en el cromosoma Y solo se pueden encontrar en machos.
Herencia ligada a X
Hay muchos más rasgos ligados al X que los rasgos ligados a Y porque el cromosoma Y es mucho más corto y menos genes que el cromosoma X.
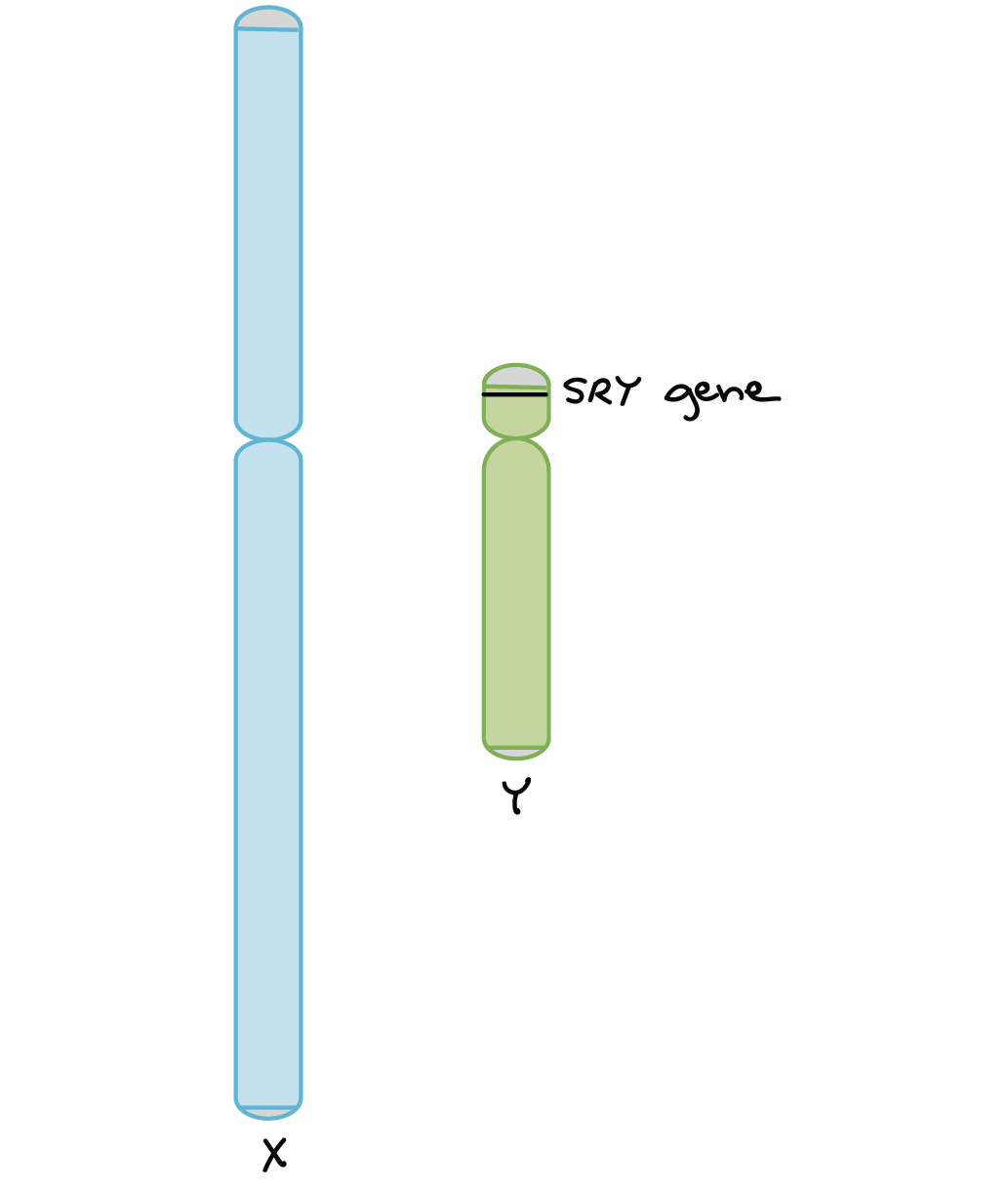
Los genes ligados al X tienen patrones de herencia distintivos porque están presentes en diferentes números en hembras (XX) y machos (XY).
Las hembras tienen dos cromosomas X, por lo que tendrá dos copias de cada gen ligado al X, lo que le dará la oportunidad de ser homocigótica o heterocigótica para cada gen ligado al sexo.
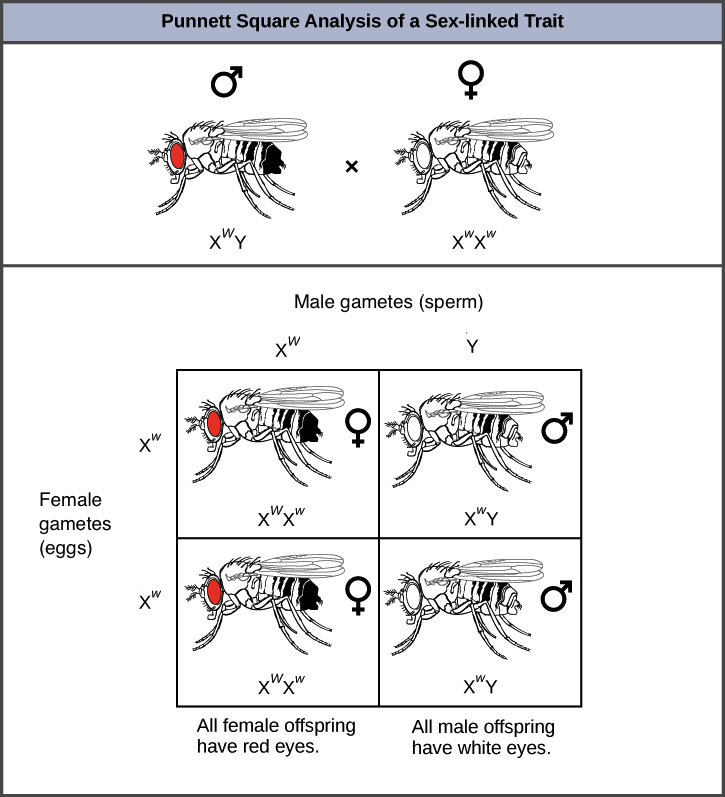
Trastornos ligados al X
Los trastornos genéticos humanos ligados al X son mucho más comunes en hombres que en mujeres. Dado que los machos solo tienen un cromosoma X, y por lo tanto una copia de cualquier gen ligado al X, se expresará cualquier alelo que herede el macho para un gen ligado al X.
Un ejemplo de ello es el trastorno de coagulación sanguínea, la hemofilia. Las mujeres heterocigóticas para la hemofilia son portadoras, y por lo general no muestran ningún síntoma por sí mismas.
Los hijos de estas mujeres tienen un 5 50, por ciento de probabilidad de tener hemofilia. Las hijas tienen pocas posibilidades de tener hemofilia (a menos que el padre también la tenga), y en cambio tendrán un 5 50, por ciento de probabilidad de ser portadoras.
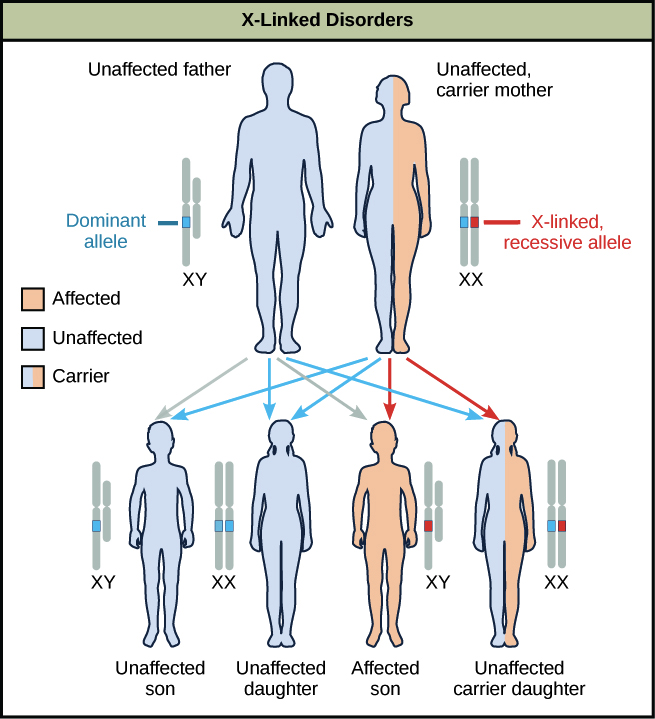
X-inactivación
Si los machos pueden sobrevivir con un solo cromosoma X, ¿por qué no causa problemas a las mujeres que tienen dos cromosomas X?
Al final resulta que, para las hembras, la mayoría de los genes en uno de los cromosomas X se inactiva, formando un cuerpo Barr. Esta inactivación ocurre aleatoriamente durante el desarrollo embrionario.
Ejemplo:
Un ejemplo común de inactivación de X se observa en gatos. Si una gata hembra es heterocigota para los alelos negros y bronceados de un gen de color de pelaje que se encuentra en la X, dos Xs (y por lo tanto, los dos alelos del gen del color del pelaje) se inactivarán al azar en diferentes células durante el desarrollo.
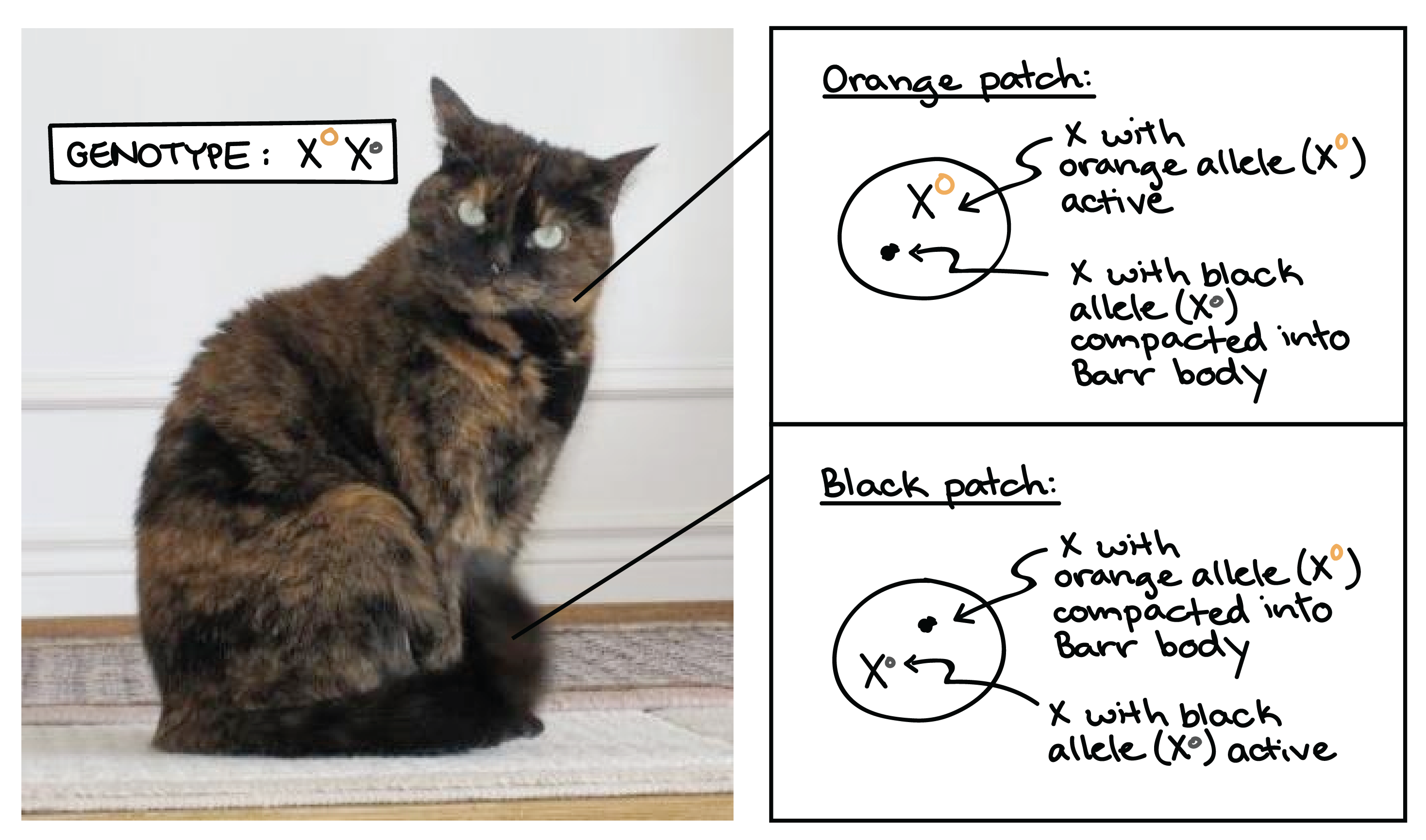
El resultado de esto es un patrón de abrigo de carey, conformado por parches alternados de pelaje negro y bronceado.
Aneuploidía cromosómica sexual
La aneuploidía, o trastornos del número de cromosomas, generalmente son causados por la no disyunción. Esto ocurre cuando los pares de cromosomas homólogos o cromátidas hermanas no logran separarse durante la división celular.

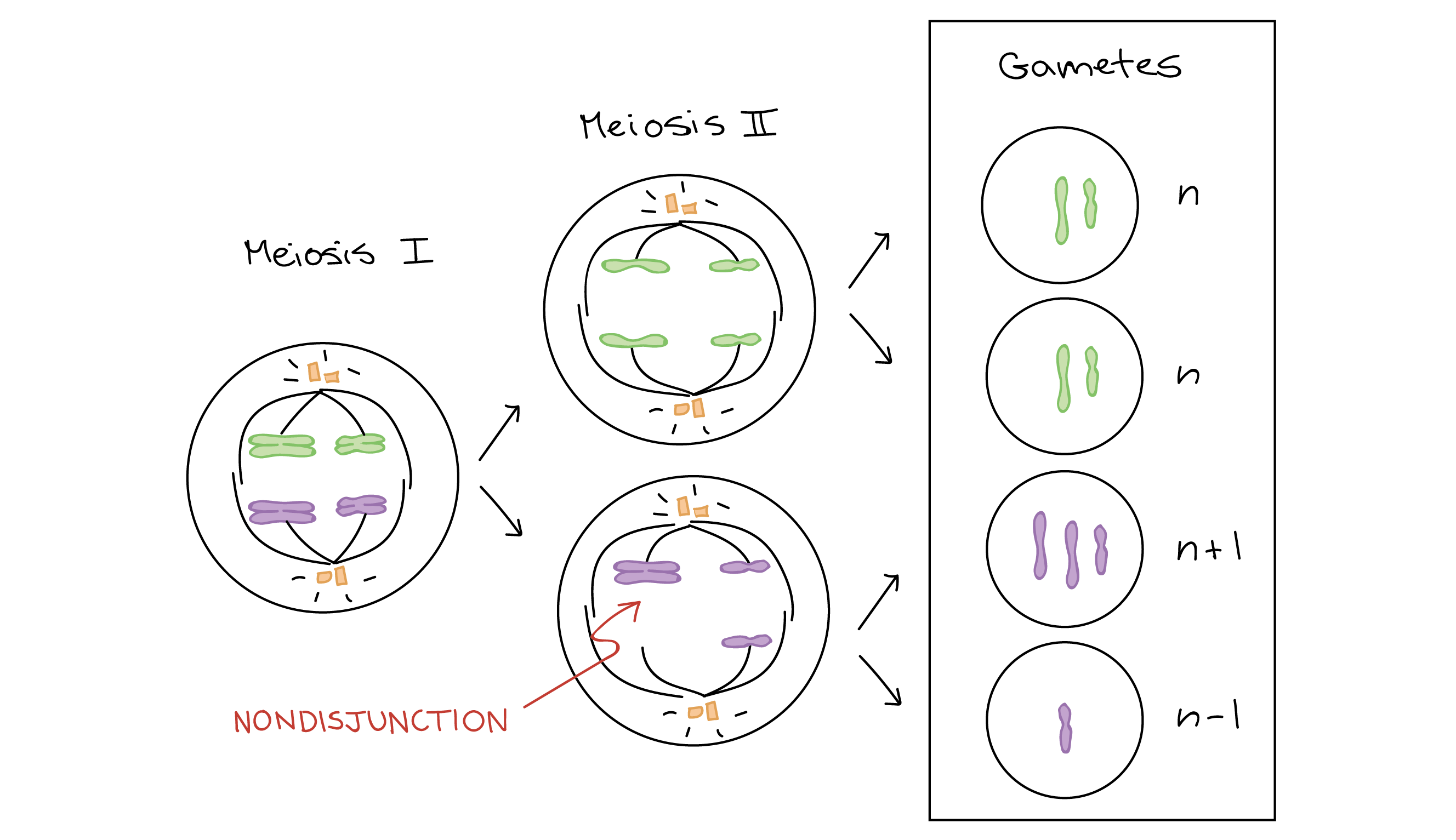
Los individuos que tienen aneuploidía autosómica rara vez sobreviven hasta el nacimiento. Sin embargo, debido al tamaño del cromosoma X y debido a la inactivación del cromosoma X, las aneuploidías del cromosoma X tienden a ser mucho menos dañinas.

En el síndrome de Klinefelter, los machos tienen uno o más cromosomas X adicionales, lo que lleva a un genotipo de XXY. (¡O en casos raros, XXXY o XXXXY!) Los hombres afectados pueden ser infértiles o desarrollar vello corporal y facial menos denso que otros hombres.
Las mujeres afectadas con el síndrome Triple X tienen un genotipo XXX. Las mujeres con síndrome Triple X tienen características sexuales femeninas y son fértiles (capaces de tener hijos).
Las mujeres con síndrome de Turner carecen de parte o la totalidad de uno de sus cromosomas X (dejándola con una sola X funcional). Las personas con este trastorno se desarrollan como mujeres, pero a menudo tienen baja estatura y pueden experimentar infertilidad y dificultades de aprendizaje.
Errores comunes y conceptos erróneos
-
Algunas personas piensan que un rasgo recesivo ligado al X aparecerá con mayor frecuencia en las mujeres porque tienen dos cromosomas X. Sin embargo, las mujeres tienen menos probabilidades de expresar rasgos recesivos ligados al X porque existe la posibilidad de que un alelo “bueno” enmascara un alelo “malo”. Por otra parte, si un macho recibe un alelo “malo” de su madre, no tiene ninguna posibilidad de obtener un alelo “bueno” de su padre (que proporciona una Y) para ocultar el malo.
-
La codominancia y la inactivación de X no son lo mismo. Aunque estos dos conceptos pueden resultar en organismos de aspecto similar, un individuo heterocigótico que expresa un rasgo codominante expresará ambos alelos total y separadamente.En la inactivación de X, las hembras expresan solo un cromosoma X en cada célula, lo que significa que los genes en el cromosoma X se expresan individualmente en lugar de en un par. Debido a que el cromosoma X inactivado no es el mismo en todas las células, las células vecinas pueden expresar diferentes proteínas si diferentes cromosomas X portan diferentes alelos.