
4: Bacterias - Paredes Celulares
- Page ID
- 56003
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Es importante señalar que no todas las bacterias tienen una pared celular. Dicho esto, sin embargo, también es importante señalar que la mayoría de las bacterias (alrededor del 90%) tienen una pared celular y suelen tener uno de dos tipos: una pared celular gram positiva o una pared celular gram negativa. Los dos tipos diferentes de pared celular se pueden identificar en el laboratorio mediante una tinción diferencial conocida como tinción Gram. Desarrollado en 1884, ha estado en uso desde entonces. Originalmente, no se sabía por qué la tinción de Gram permitía una separación tan confiable de bacterias en dos grupos. Una vez inventado el microscopio electrónico en la década de 1940, se encontró que la diferencia de tinción se correlacionaba con diferencias en las paredes celulares. Aquí hay un sitio web que muestra los pasos reales de la mancha Gram. Después de aplicar esta técnica de tinción las bacterias gram positivas se mancharán de púrpura, mientras que las bacterias gram negativas se mancharán de rosa.

Descripción general de las paredes celulares bacterianas
Una pared celular, no sólo de bacterias sino para todos los organismos, se encuentra fuera de la membrana celular. Es una capa adicional que normalmente proporciona cierta resistencia de la que carece la membrana celular, al tener una estructura semirrígida.
Tanto las paredes celulares gram positivas como las gram negativas contienen un ingrediente conocido como peptidoglicano (también conocido como mureína). Esta sustancia en particular no se ha encontrado en ningún otro lugar de la Tierra, aparte de las paredes celulares de las bacterias. Pero ambos tipos de pared celular bacteriana también contienen ingredientes adicionales, lo que hace que la pared celular bacteriana sea una estructura compleja en general, particularmente cuando se compara con las paredes celulares de microbios eucariotas. Las paredes celulares de los microbios eucariotas suelen estar compuestas por un solo ingrediente, como la celulosa que se encuentra en las paredes celulares de algas o la quitina en las paredes celulares fúngicas.
La pared celular bacteriana también realiza varias funciones, además de proporcionar fuerza general a la célula. También ayuda a mantener la forma celular, lo cual es importante para la forma en que la célula crecerá, se reproducirá, obtendrá nutrientes y se moverá. Protege a la célula de la lisis osmótica, ya que la célula se mueve de un ambiente a otro o transporta en nutrientes de su entorno. Dado que el agua puede moverse libremente tanto a través de la membrana celular como de la pared celular, la célula está en riesgo de un desequilibrio osmótico, lo que podría ejercer presión sobre la membrana plasmática relativamente débil. Los estudios han demostrado en realidad que la presión interna de una celda es similar a la presión que se encuentra dentro de una llanta de automóvil completamente inflada. ¡Eso es mucha presión para que la membrana plasmática resista! La pared celular puede mantener alejadas ciertas moléculas, como las toxinas, particularmente para las bacterias gram negativas. Y por último, la pared celular bacteriana puede contribuir a la patogenicidad o enfermedad —causando capacidad de la célula para ciertos patógenos bacterianos.
Estructura del peptidoglicano
Empecemos por peptidoglicano, ya que es un ingrediente que ambas paredes celulares bacterianas tienen en común. El peptidoglicano es un polisacárido compuesto por dos derivados de glucosa, N-acetilglucosamina (NAG) y ácido N-acetilmurámico (NAM), alternados en cadenas largas. Las cadenas están reticuladas entre sí por un tetrapéptido que se extiende fuera de la unidad de azúcar NAM, permitiendo que se forme una estructura en forma de celosía. Los cuatro aminoácidos que componen el tetrapéptido son: L-alanina, D-glutamina, L-lisina o ácido meso-diaminopimélico (DPA) y D-alanina. Típicamente, solo la forma L-isomérica de aminoácidos es utilizada por las células, pero el uso de los D-aminoácidos de imagen especular proporciona protección contra proteasas que podrían comprometer la integridad de la pared celular atacando el peptidoglicano. Los tetrapéptidos se pueden reticular directamente entre sí, con la D-alanina en un tetrapéptido uniéndose a la L-lisina/DPA en otro tetrapéptido. En muchas bacterias gram positivas hay un puente cruzado de cinco aminoácidos como la glicina (interpuente peptídico) que sirve para conectar un tetrapéptido a otro. En cualquier caso, la reticulación sirve para aumentar la resistencia de la estructura general, con mayor resistencia derivada de la reticulación completa, donde cada tetrapéptido está unido de alguna manera a un tetrapéptido en otra cadena NAG-NAM.
Si bien aún se desconoce mucho sobre el peptidoglicano, la investigación realizada en los últimos diez años sugiere que el peptidoglicano se sintetiza como un cilindro con una subestructura enrollada, donde cada bobina se reticula con la bobina que está junto a ella, creando una estructura aún más fuerte en general.
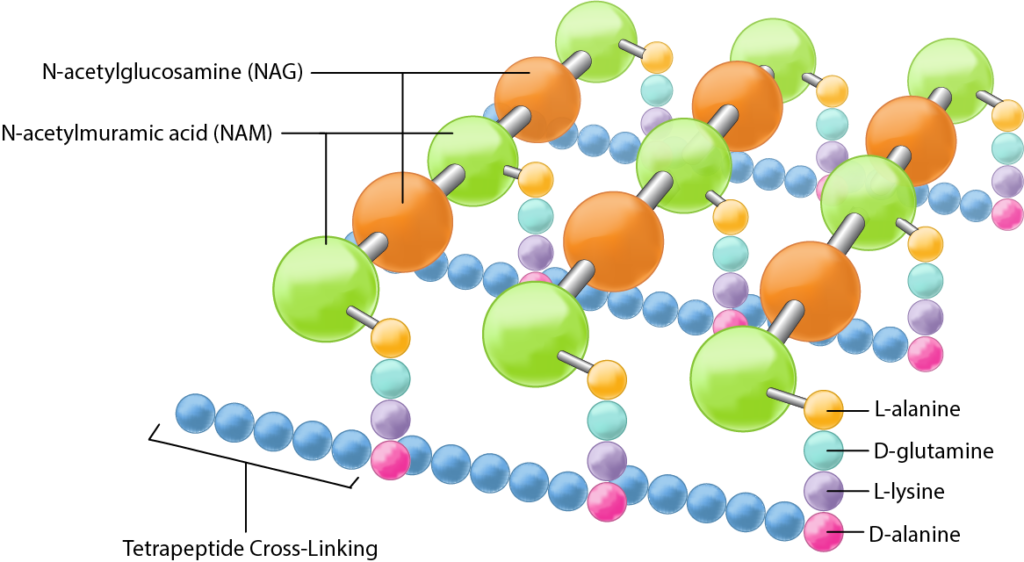
Estructura del peptidoglicano.
Paredes celulares Gram Positivas
Las paredes celulares de las bacterias gram positivas están compuestas predominantemente por peptidoglicano. De hecho, el peptidoglicano puede representar hasta el 90% de la pared celular, formándose capa tras capa alrededor de la membrana celular. Los tetrapéptidos NAM están típicamente reticulados con un interpuente peptídico y la reticulación completa es común. Todo esto se combina para crear una pared celular increíblemente fuerte.
El componente adicional en una pared celular gram positiva es el ácido teicoico, un glicopolímero, que está incrustado dentro de las capas de peptidoglicano. Se cree que el ácido teicoico juega varios papeles importantes para la célula, como la generación de la carga negativa neta de la célula, que es esencial para el desarrollo de una fuerza motriz protónica. El ácido teicoico contribuye a la rigidez general de la pared celular, lo cual es importante para el mantenimiento de la forma celular, particularmente en organismos en forma de varilla. También hay evidencia de que los ácidos teicoicos participan en la división celular, al interactuar con la maquinaria de biosíntesis de peptidoglicanos. Por último, los ácidos teicoicos parecen desempeñar un papel en la resistencia a condiciones adversas como altas temperaturas y altas concentraciones de sal, así como a los antibióticos β-lactámicos. Los ácidos teicoicos pueden unirse covalentemente a peptidoglicano (ácidos teicoicos de pared o WTA) o conectarse a la membrana celular a través de un anclaje lipídico, en cuyo caso se le conoce como ácido lipoteicoico.
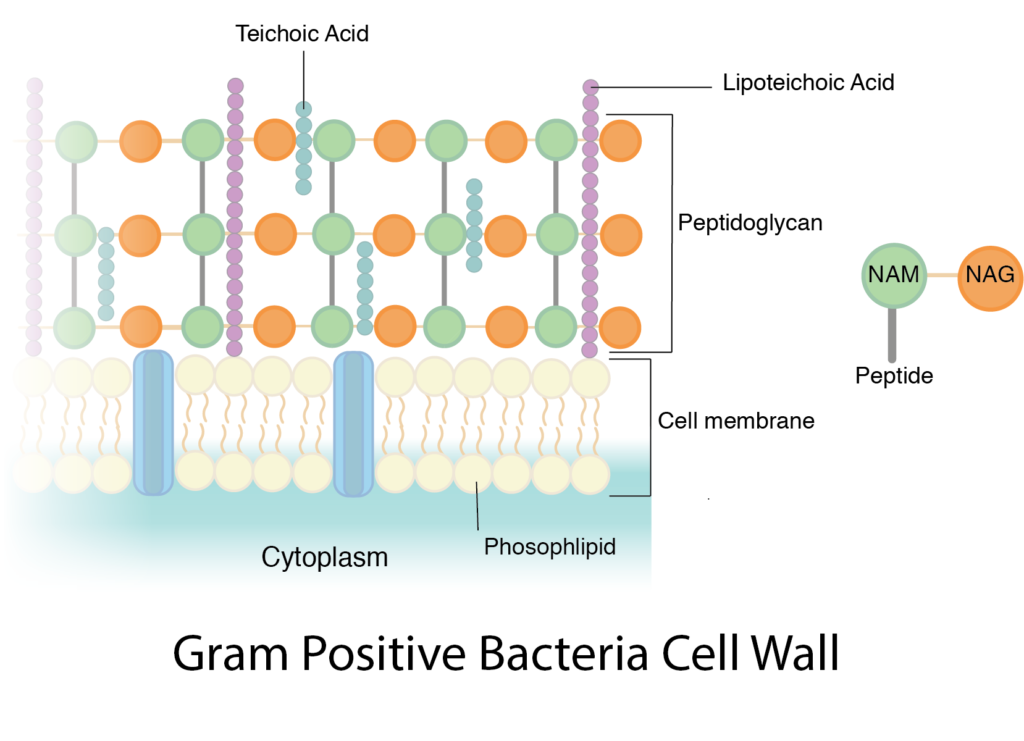
Dado que el peptidoglicano es relativamente poroso, la mayoría de las sustancias pueden pasar a través de la pared celular gram positiva con poca dificultad. Pero algunos nutrientes son demasiado grandes, requiriendo que la célula confíe en el uso de exoenzimas. Estas enzimas extracelulares se producen dentro del citoplasma de la célula y luego se secretan más allá de la membrana celular, a través de la pared celular, donde funcionan fuera de la célula para descomponer grandes macromoléculas en componentes más pequeños.
Paredes celulares Gram negativas
Las paredes celulares de las bacterias gram negativas son más complejas que las de las bacterias gram positivas, con más ingredientes en general. También contienen peptidoglicano, aunque solo un par de capas, lo que representa 5-10% de la pared celular total. Lo más notable de la pared celular gram negativa es la presencia de una membrana plasmática ubicada fuera de las capas de peptidoglicano, conocida como membrana externa. Esto conforma la mayor parte de la pared celular gram negativa. La membrana externa está compuesta por una bicapa lipídica, muy similar en composición a la membrana celular con cabezas polares, colas de ácidos grasos y proteínas integrales. Se diferencia de la membrana celular por la presencia de moléculas grandes conocidas como lipopolisacárido (LPS), las cuales se anclan a la membrana externa y se proyectan desde la célula hacia el ambiente. El LPS se compone de tres componentes diferentes: 1) el O-antígeno u O-polisacárido, que representa la parte más externa de la estructura, 2) el polisacárido central y 3) el lípido A, que ancla el LPS en la membrana externa. Se sabe que el LPS cumple muchas funciones diferentes para la célula, como contribuir a la carga negativa neta para la célula, ayudar a estabilizar la membrana externa y proporcionar protección contra ciertas sustancias químicas bloqueando físicamente el acceso a otras partes de la pared celular. Además, el LPS juega un papel en la respuesta del hospedador a bacterias gramnegativas patógenas. El antígeno O desencadena una respuesta inmune en un huésped infectado, provocando la generación de antibióticos específicos para esa parte del LPS (piense en E. coli O 157). El lípido A actúa como toxina, específicamente una endotoxina, causando síntomas generales de enfermedad como fiebre y diarrea. Una gran cantidad de lípido A liberado en el torrente sanguíneo puede desencadenar un shock endotóxico, una respuesta inflamatoria en todo el cuerpo que puede poner en peligro la vida.
La membrana externa sí presenta un obstáculo para la célula. Si bien hay ciertas moléculas que le gustaría mantener fuera, como los antibióticos y los químicos tóxicos, hay nutrientes que le gustaría dejar entrar y la bicapa lipídica adicional presenta una barrera formidable. Las moléculas grandes son descompuestas por enzimas, con el fin de permitirles pasar el LPS. En lugar de exoenzimas (como las bacterias gram positivas), las bacterias gram negativas utilizan enzimas periplásmicas que se almacenan en el periplasma. ¿Dónde está el periplasma, usted pregunta? Es el espacio ubicado entre la superficie externa de la membrana celular y la superficie interna de la membrana externa, y contiene el peptidoglicano gram negativo. Una vez que las enzimas periplásmicas han descompuesto los nutrientes en moléculas más pequeñas que pueden pasar del LPS, aún necesitan ser transportadas a través de la membrana externa, específicamente la bicapa lipídica. Las células Gram negativas utilizan porinas, que son proteínas transmembrana compuestas por un trímero de tres subunidades, que forman un poro a través de la membrana. Algunas porinas son inespecíficas y transportan cualquier molécula que se ajuste, mientras que algunas porinas son específicas y solo transportan sustancias que reconocen mediante el uso de un sitio de unión. Una vez a través de la membrana externa y en el periplasma, las moléculas se abren paso a través de las capas porosas de peptidoglicano antes de ser transportadas por proteínas integrales a través de la membrana celular.
Las capas de peptidoglicano están unidas a la membrana externa mediante el uso de una lipoproteína conocida como lipoproteína de Braun (buen ol' Dr. Braun). En un extremo, la lipoproteína se une covalentemente al peptidoglicano mientras que el otro extremo está incrustado en la membrana externa a través de su cabeza polar. Este enlace entre las dos capas proporciona integridad estructural y resistencia adicionales.
Bacterias inusuales y sin pared
Habiendo enfatizado la importancia de una pared celular y el ingrediente peptidoglicano tanto para la bacteria gram positiva como para la gram negativa, parece importante señalar algunas excepciones también. Las bacterias pertenecientes al filo Chlamydiae parecen carecer de peptidoglicano, aunque sus paredes celulares tienen una estructura gram negativa en todos los demás aspectos (es decir, membrana externa, LPS, porina, etc.). Se ha sugerido que podrían estar usando una capa proteica que funciona de la misma manera que el peptidoglicano. Esto tiene una ventaja para la célula en proporcionar resistencia a los antibióticos β-lactámicos (como la penicilina), que atacan al peptidoglicano.
Las bacterias pertenecientes al filo Tenericutes carecen de una pared celular por completo, lo que las hace extremadamente susceptibles a los cambios osmóticos. A menudo fortalecen algo su membrana celular mediante la adición de esteroles, una sustancia generalmente asociada a las membranas celulares eucariotas. Muchos miembros de este filo son patógenos, eligiendo esconderse dentro del ambiente protector de un huésped.
Palabras clave
pared celular, bacterias gram positivas, bacterias gram negativas, tinción Gram, peptidoglicano, mureína, lisis osmótica, N-acetilglucosamina (NAG), ácido N-acetilmurámico (NAM), tetrapéptido, L-alanina, D-glutamina, L-lisina, ácido meso-diaminopimélico (DPA), D-alanina, entrecruzamiento directo, interpuente peptídico, reticulación completa, ácido teicoico, ácido teicoico de pared (WTA), ácido lipoteicoico, exoenzimas, membrana externa, lipopolisacárido (LPS), O-antígeno u O-polisacárido, polisacárido central, lípido A, endotoxina, enzimas periplásmicas, periplasma, porinas, lipoproteína de Braun, Chlamydiae, Tenericutes, esteroles.
Preguntas/objetivos esenciales
- ¿Cuáles son las características y funciones básicas de la pared celular en Bacterias?
- ¿Qué es la tinción Gram y cómo se relaciona con los diferentes tipos de bacterias de la pared celular?
- ¿Cuál es la estructura unitaria básica del peptidoglicano? ¿Qué componentes están presentes y cómo interactúan? Ser capaz de diagrama peptidoglicano y sus componentes.
- ¿Qué es el entrecruzamiento y por qué juega este papel tan importante en la pared celular? ¿Qué diferentes tipos de reticulación hay?
- ¿Por qué los D-aminoácidos son inusuales y cómo tener D-aminoácidos en el peptidoglicano mantiene estable esta macromolécula?
- ¿Cuáles son las diferencias entre los organismos gram positivos y negativos en términos de grosor del peptidoglicano, diferentes constituyentes de PG y variaciones en la reticulación y fuerza, y otras moléculas asociadas con la pared celular?
- ¿Qué es el ácido teicoico y cuáles son sus roles y funciones propuestos? ¿Qué son los ácidos lipteicoicos?
- ¿Cuál es el periplasma de las bacterias gram negativas? ¿Qué propósito puede servir? ¿Qué alternativas hay disponibles para las células?
- ¿Cuál es la composición general de la membrana externa de los microorganismos gramnegativos, su función y propiedades tóxicas? ¿Cómo se vincula a la célula? ¿Qué es una porina y cuáles son sus funciones?
- ¿Qué grupo de bacterias carecen de peptidoglicano en su pared celular? ¿Qué ventaja confiere esto?
- ¿Qué grupo de bacterias normalmente no tiene paredes celulares y cómo se mantienen?
Preguntas Exploratorias (OPCIONAL)
- ¿Cómo se relaciona el mecanismo de la tinción de Gram con componentes específicos de la pared celular bacteriana?