
5: Bacterias - Componentes Internos
- Page ID
- 56076
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Ya hemos cubierto los principales componentes internos que se encuentran en todas las bacterias, es decir, el citoplasma, el nucleoide y los ribosomas. Recuerde que generalmente se piensa que las bacterias carecen de orgánulos, esos compartimentos ligados a la membrana bilipídica tan prevalentes en las células eucariotas (aunque algunos científicos argumentan que las bacterias poseen estructuras que podrían pensarse como simples orgánulos). Pero las bacterias pueden ser más complejas, con una variedad de componentes internos adicionales por encontrar que pueden contribuir a sus capacidades. La mayoría de estos componentes son citoplásmicos pero algunos de ellos son periplásmicos, localizados en el espacio entre la membrana citoplasmática y externa en bacterias gram negativas.
Citoesqueleto
Originalmente se pensó que las bacterias carecían de un citoesqueleto, un componente significativo de las células eucariotas. En los últimos 20 años, sin embargo, los científicos han descubierto filamentos bacterianos hechos de proteínas que son análogos a las proteínas citoesqueléticas que se encuentran en los eucariotas. También se ha determinado que el citoesqueleto bacteriano juega un papel importante en la forma celular, división celular e integridad de la pared celular.
FtsZ
FtsZ, homóloga a la proteína eucariota tubulina, forma una estructura de anillo en el centro de la célula durante la división celular, atrayendo otras proteínas al área para construir un tabique que eventualmente separará las dos células hijas resultantes.
MReb
MReB, homólogo a la proteína eucariota actina, se encuentra en bacilo y bacterias en forma de espiral y juega un papel esencial en la formación de la forma celular. MReb asume una configuración helicoidal que recorre la longitud de la célula y dicta las actividades de la maquinaria de síntesis de peptidoglicanos, asegurando una forma no esférica.
Crescentin
La crescentina, homóloga a las proteínas eucariotas laminas y queratina, se encuentra en bacterias en forma de espiral con una sola curva. La proteína se ensambla longitudinalmente en la curvatura interna de la célula, doblando la célula en su forma final.
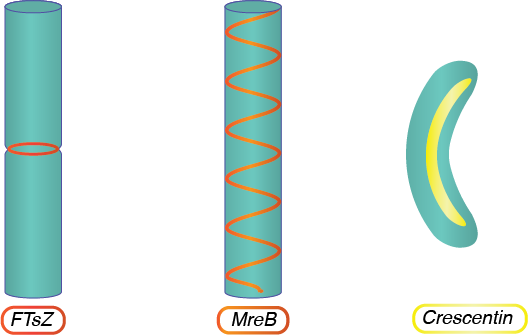
Estructuras del citoesqueleto.
Inclusiones
Las inclusiones bacterianas se definen generalmente como una estructura distinta localizada dentro del citoplasma o periplasma de la célula. Pueden variar en complejidad, desde una simple compilación de químicos como los cristales, hasta estructuras bastante complejas que comienzan a rivalizar con la de los orgánulos eucariotas, completos con una capa externa membranosa. Su papel suele ser almacenar componentes como reservas metabólicas para la célula cuando una sustancia se encuentra en exceso, pero también pueden desempeñar un papel en la motilidad y las funciones metabólicas también.
Almacenamiento de carbono
El carbono es la sustancia más común para ser almacenada por una célula, ya que todas las células son a base de carbono. Además, los compuestos de carbono a menudo pueden ser desagregados rápidamente por la célula, por lo que también pueden servir como fuentes de energía. Una de las inclusiones más simples y comunes para el almacenamiento de carbono es el glucógeno, en el que las unidades de glucosa están unidas entre sí en una estructura de polisacárido multiramificante.
Otra forma común para que las bacterias almacenen carbono es en forma de poli-β-hidroxibutirato (PHB), un gránulo que se forma cuando las unidades de ácido β-hidroxibutírico se agregan juntas. Este lípido es muy parecido al plástico en su composición, lo que lleva a algunos científicos a investigar la posibilidad de utilizarlos como plástico biodegradable. Los gránulos de PHB en realidad tienen una cubierta compuesta tanto por proteína como por una pequeña cantidad de fosfolípido. Tanto el glucógeno como el PHB se forman cuando hay un exceso de carbono y luego se descomponen por la célula posteriormente tanto para carbono como para energía.
Almacenamiento inorgánico
A menudo las bacterias necesitan algo más que el carbono, ya sea para la síntesis de componentes celulares o como reserva de energía alternativa. Los gránulos de polifosfato permiten la acumulación de fosfato inorgánico (PO43-), donde el fosfato puede ser utilizado para producir ácido nucleico (¿recuerdas la cadena principal de azúcar-fosfato?) o ATP (adenosina trifosfato, por supuesto).
Otras células necesitan azufre como fuente de electrones para su metabolismo y almacenarán el exceso de azufre en forma de glóbulos de azufre, lo que resulta cuando la célula oxida sulfuro de hidrógeno (H2S) a azufre elemental (S0), resultando en la formación de inclusiones refráctiles.
Funciones que no son de almacenamiento
Hay momentos en que una bacteria necesita hacer algo más allá del simple almacenamiento de compuestos orgánicos o inorgánicos para su uso en el metabolismo y hay inclusiones para ayudar con estas funciones de no almacenamiento. Un ejemplo de ello son las vacuolas de gas, las cuales son utilizadas por la celda para controlar la flotabilidad en una columna de agua, proporcionando a la celda cierto control sobre dónde se encuentra en el ambiente. Es una forma limitada de motilidad, solo en el eje vertical. Las vacuolas de gas están compuestas por conglomerados de vesículas de gas, estructuras cilíndricas que son a la vez huecas y rígidas. Las vesículas de gas son libremente permeables a todo tipo de gases por difusión pasiva y pueden construirse o colapsarse rápidamente, según sea necesario por la célula para ascender o descender.
Los magnetosomas son inclusiones que contienen largas cadenas de magnetita (Fe3O4), las cuales son utilizadas por la celda como brújula en campos geomagnéticos, para la orientación dentro de su entorno. Las bacterias magnetotácticas son típicamente microaerofílicas, prefiriendo un ambiente con un nivel de oxígeno más bajo que la atmósfera. El magenetosoma permite que las células localicen la profundidad óptima para su crecimiento. Los magenetosomas tienen una verdadera bicapa lipídica, que recuerda a los orgánulos eucariotas, pero en realidad es una invaginación de la membrana plasmática de la célula que ha sido modificada con proteínas específicas.
Microcompartimientos
Los microcompartimentos bacterianos (BMC) son únicos de otras inclusiones en virtud de su estructura y funcionalidad. Tienen forma icosaédrica y están compuestas por una cubierta proteica compuesta por diversas proteínas de la familia BMC. Si bien su papel exacto varía, todos participan en funciones más allá del simple almacenamiento de sustancias. Estos compartimentos proporcionan tanto una ubicación como las sustancias (generalmente enzimas) necesarias para actividades metabólicas particulares.
El ejemplo mejor estudiado de una BMC es el carboxisoma, que se encuentran en muchas bacterias fijadoras de CO2. Los carboxisomas contienen la enzima ribulosa-1,5-bisfosfato carboxilasa (afortunadamente también se conoce como RubiSco), que juega un papel crucial en la conversión del CO2 en azúcar. El carboxisoma también juega un papel en la concentración de CO2, asegurando así que los componentes necesarios para la fijación de CO2 estén todos en el mismo lugar al mismo tiempo.
Anammoxosoma
El anammoxosoma es un gran compartimento unido a membrana que se encuentra en células bacterianas capaces de llevar a cabo la reacción annamox (una idación aeróbica de am monium ox), donde el amonio (NH4+) y el nitrito (NO2-) se convierten en gas dinitrógeno (N2). El proceso se realiza como una forma para que la célula obtenga energía, utilizando el amonio como donante de electrones y el nitrito como aceptación de electrones, con la producción resultante de gas nitrógeno. Esta conversión química del nitrógeno es importante para el ciclo del nitrógeno.
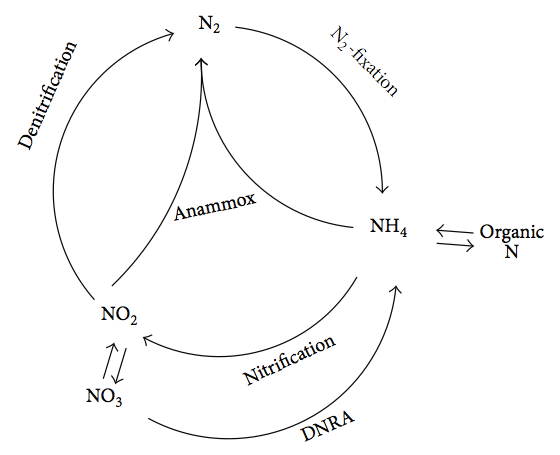
Ciclo de Nitrógeno. Por Shou-Qing Ni y Jian Zhang [CC BY 3.0], vía Wikimedia Commons
Clorosoma
Encontrado en algunas bacterias fototróficas, un clorosoma es una estructura altamente eficiente para capturar intensidades de poca luz. Al revestir el perímetro interior de la membrana celular, cada clorosoma puede contener hasta 250,000 moléculas de bacterioclorofila, dispuestas en matrices densas. La luz cosechada se transfiere a los centros de reacción en la membrana celular, permitiendo la conversión de energía luminosa a energía química en forma de ATP. El clorosoma está limitado por una monocapa lipídica.
Plasmido
Un plásmido es una pieza extracromosómica de ADN que algunas bacterias tienen, además del material genético que se encuentra en el nucleoide. Está compuesto por ADN bicatenario y es típicamente circular, aunque se han encontrado plásmidos lineales. Los plásmidos se describen como “no esenciales” para la célula, donde la célula puede funcionar normalmente en su ausencia. Pero si bien los plásmidos solo tienen unos pocos genes, pueden conferir capacidades importantes para la célula, como la resistencia a antibióticos. Los plásmidos se replican independientemente de la célula y pueden perderse (conocido como curado), ya sea espontáneamente o debido a la exposición a condiciones adversas, como la luz UV, la inanición de timina o el crecimiento por encima de las condiciones óptimas. Algunos plásmidos, conocidos como episomas, pueden integrarse en el cromosoma celular donde los genes se replicarán durante la división celular.
Endospore
Luego está la endospora, una maravilla de la ingeniería bacteriana. Esta se encuentra bajo el encabezamiento “componentes internos bacterianos”, pero es importante señalar que una endospora no es una estructura interna o externa tanto como una conversión de la célula en una forma alternativa. Las células empiezan como una célula vegetativa, haciendo todas las cosas que se supone que debe hacer una célula (metabolizar, reproducir, cortar el césped...). Si se exponen a condiciones hostiles (desización, calor alto, un vecino enojado...) y tienen la habilidad, podrían convertirse de célula vegetativa en endospora. La endospora se forma realmente dentro de la célula vegetativa (¿eso no la convierte en una estructura interna?) y luego la célula vegetativa se lisa, liberando la endospora (¿eso la convierte en una estructura externa?).
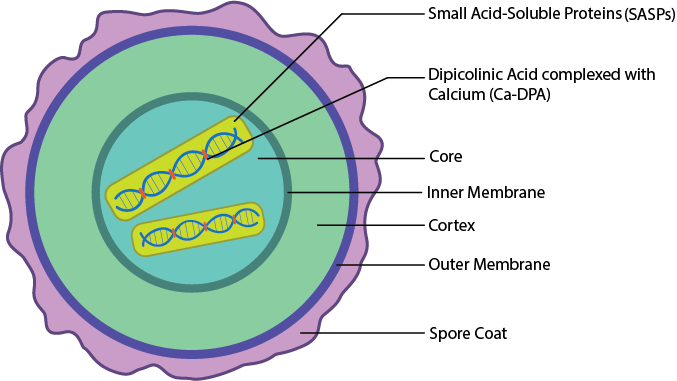
Capas de Endospore.
Las endosporas solo están formadas por unos pocos géneros gram positivos y proporcionan a la célula resistencia a una amplia variedad de condiciones duras, como inanición, temperaturas extremas, exposición al secado, luz UV, productos químicos, enzimas y radiación. Si bien la célula vegetativa es la forma activa para las células bacterianas (en crecimiento, metabolización, etc.), la endospora puede pensarse como una forma latente de la célula. Permite la supervivencia de condiciones adversas, pero no permite que la célula crezca o se reproduzca.
Estructura
Para ser tan increíblemente resistentes a tantas sustancias y condiciones ambientales diferentes, son necesarias muchas capas diferentes. La endospora bacteriana tiene muchas capas diferentes, comenzando con un núcleo en el centro. El núcleo es la ubicación del nucleoide, los ribosomas y el citoplasma de la célula, en una forma extremadamente deshidratada. Por lo general, contiene solo el 25% del agua que se encuentra en una celda vegetativa, aumentando la resistencia al calor. El ADN está protegido además por la presencia de pequeñas proteínas solubles en ácido (SASP), que estabilizan el ADN y lo protegen de la degradación. La estabilización del ADN se incrementa por la presencia de ácido dipicolínico complejado con calcio (Ca-dPA), que se inserta entre las bases de ADN. El núcleo está envuelto en una membrana interna que proporciona una barrera de permeabilidad a los productos químicos, que luego es rodeada por la corteza, una capa gruesa que consiste en peptidoglicano con menos reticulación que la que se encuentra en la célula vegetativa. La corteza está envuelta en una membrana externa. Por último, se encuentran varias capas de esporas hechas de proteína, que brindan protección contra el estrés ambiental como químicos y enzimas.
Esporulación: conversión de célula vegetativa a endospora
La esporulación, la conversión de la célula vegetativa en la endospora altamente protectora, suele ocurrir cuando la supervivencia de la célula se ve amenazada de alguna manera. El proceso real es muy complejo y suele tardar varias horas hasta su finalización. Inicialmente las células esporulantes replican su ADN, como si estuviera a punto de sufrir división celular. Un tabique se forma asimétricamente, secuestrando una copia del cromosoma en un extremo de la célula (llamada la forespora). Se produce la síntesis de sustancias específicas de endospora, alterando la forespora y dando lugar al desarrollo de las capas específicas para una endospora, así como a la deshidratación. Finalmente, la “célula madre” es lisada, permitiendo la liberación de la endospora madura al ambiente.
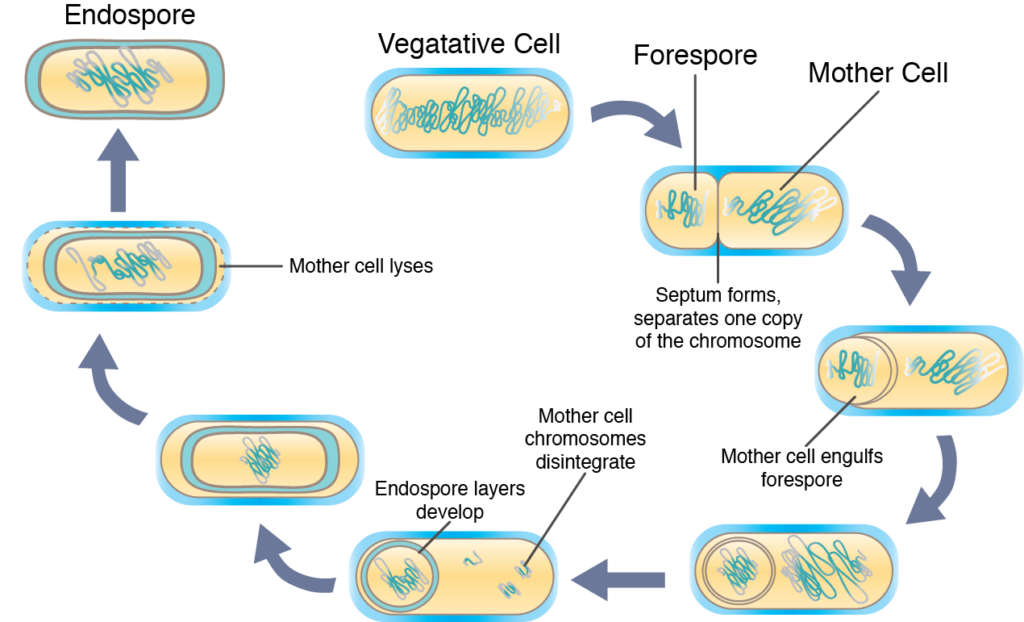
Esporulación.
Conversión de endospora a célula vegetativa
La endospora permanece latente hasta que las condiciones ambientales mejoran, provocando un cambio químico que inicia la expresión génica. Hay tres etapas distintas en la conversión de una endospora a las células vegetativas metabólicamente activas: 1) activación, etapa de preparación que puede iniciarse por la aplicación de calor; 2) germinación, cuando la endospora se vuelve metabólicamente activa y comienza a tomar agua; 3) excrecencia, cuando la célula vegetativa emerge completamente de la cáscara de endospora.
Palabras clave
citoesqueleto, FtsZ, tubulina, MReB, actina, crescentina, lamina, queratina, inclusión, glucógeno, poli-β-hidroxibutirato (PHB), gránulo de polifosfato, glóbulo de azufre, vacuola de gas, vesícula gaseosa, magnetosoma, microaerofílico, microcompartimento, microcompartimentos bacterianos (BMC), carboxisoma, ribulosa-1,5-bisfosfato carboxilasa, RubisCO, anammoxosoma, reacción annamox, clorosoma, plásmido, curado, episoma, endospora, célula vegetativa, núcleo, pequeñas proteínas solubles en ácido (SASPs), ácido dipicolínico, Ca-dPA, membrana interna, corteza, membrana externa, recubrimiento de esporas, esporulación, preporo, activación, germinación, excrecencia.
Preguntas/Objetivos Esenciales
- ¿Cuáles son los papeles y la composición del citoesqueleto bacteriano? ¿En qué se diferencia del citoesqueleto eucariota? ¿Cuáles son las proteínas específicas del citoesqueleto bacteriano y qué detalles se conocen de cada una?
- ¿Cuál es el propósito de las inclusiones que se encuentran en las bacterias? ¿Cuáles son sus características?
- ¿Cuáles son los ejemplos específicos de inclusiones de almacenamiento que se encuentran en las bacterias? Ser capaz de describir cada tipo en términos de estructura y propósito.
- ¿Qué otras inclusiones se encuentran en las bacterias? Ser capaz de describir cada tipo en términos de estructura y propósito.
- ¿En qué se diferencian los microcompartimentos de las inclusiones? ¿Cuáles son los ejemplos específicos? ¿Cuál es la composición y el propósito?
- ¿Qué son los anammoxosomas? ¿Cuál es su composición y propósito?
- ¿Qué son los plásmidos y qué características tienen? ¿Qué son los episomas? ¿Qué es el curado y qué lo causa?
- ¿Qué son las endosporas bacterianas? ¿Cuál es su propósito? ¿Qué características tienen? ¿Cuáles son las distintas capas de una endospora y qué papel juega cada capa?
Preguntas Exploratorias (OPCIONAL)
- ¿Qué estructuras bacterianas podrían ser útiles para los científicos para abordar los problemas sociales?