
11.3B: El Sistema Complemento
- Page ID
- 55222

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Objetivos de aprendizaje
- Describa brevemente cómo se activa la vía clásica del complemento.
- Describa brevemente los efectos beneficiosos de los siguientes productos de la vía del complemento:
- C5a
- C3a
- C3b
- C4b
- C3d
- C5b6789 n (MAC)
- Describa brevemente cómo se activa la vía lectina.
- Describa brevemente cómo se activa la vía alternativa del complemento.
En esta sección veremos cómo funciona el sistema de complemento del organismo para eliminar los agentes infecciosos. El sistema del complemento se refiere a una serie de más de 30 proteínas solubles preformadas que circulan en la sangre y bañan los fluidos que rodean los tejidos. Las proteínas circulan en forma inactiva, pero en respuesta al reconocimiento de componentes moleculares de microorganismos, se activan secuencialmente, trabajando en una cascada donde en la unión de una proteína promueve la unión de la siguiente proteína en la cascada. Existen 3 vías del complemento que conforman el sistema del complemento: la vía clásica del complemento, la vía lectina y la vía alternativa del complemento. Las vías difieren en la forma en que se inician y finalmente producen una enzima clave llamada C3 convertasa:
- La vía clásica del complemento se inicia por la activación de C1. C1 se activa principalmente interactuando con la porción Fc de las moléculas de anticuerpo IgG o IgM después de que se hayan unido a su antígeno específico. C1 también es capaz de unirse directamente a las superficies de algunos patógenos así como con la proteína C reactiva (PCR) que se produce durante la respuesta de fase aguda de la inmunidad innata.
- La vía lectina se activa por la interacción de carbohidratos microbianos (lectinas) con lectina de unión a manosa (MBL) o ficolinas que se encuentran en el plasma y fluidos tisulares.
- La vía alternativa del complemento se activa mediante la unión de C3b a superficies microbianas y a moléculas de anticuerpos.
Los resultados finales y los beneficios de defensa de cada vía, sin embargo, son los mismos. Todas las vías del complemento llevan a cabo 6 funciones benéficas de defensa innatas. Proteínas producidas por las vías del complemento:
- Inflamación desencadenante,
- Atraer quimiotácticamente los fagocitos al sitio de la infección,
- Promover la unión de antígenos a fagocitos (mayor unión u opsonización),
- Causar lisis de bacterias Gram-negativas, células humanas que muestran epítopos extraños y envolturas virales,
- Desempeñar un papel en la activación de linfocitos B ingenuos durante la inmunidad adaptativa, y
- Eliminar los complejos inmunes dañinos del cuerpo.
Ahora veremos cada una de estas vías del complemento y veremos cómo funcionan para proteger al cuerpo.
La vía clásica del complemento
La vía clásica del complemento se activa principalmente cuando un complejo de proteínas del complemento llamado C1 interactúa con la porción Fc de las moléculas de anticuerpo IgG o IgM después de que se hayan unido a su antígeno específico a través de su porción Fab. C1 también es capaz de unirse directamente a las superficies de algunos patógenos así como con la proteína C reactiva (PCR) que se produce durante la respuesta de fase aguda de la inmunidad innata. El complejo C1 está compuesto por tres proteínas del complemento llamadas C1q, C1r y C1s.
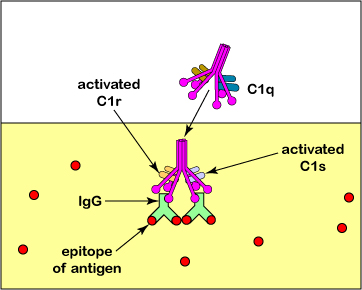
C1 también es capaz de unirse directamente a las superficies de algunos patógenos así como con la proteína C reactiva (PCR) que se produce durante la respuesta de fase aguda de la inmunidad innata.
2. La unión de C1q activa la porción C1r de C1 que, a su vez, activa C1s. Esta activación da actividad enzimática de C1s para escindir la proteína C4 del complemento en C4a y C4b (ver Figura\(\PageIndex{2}\) A y Figura\(\PageIndex{2}\) B).
3. C2 luego se une a C4b y se escinde por C1 en C2a y C2b (ver Figura\(\PageIndex{3}\) A y Figura\(\PageIndex{3}\) B).
4. C4b y C2a se combinan para formar C4B2a, la convertasa C3. La convertasa C3 ahora puede escindir cientos de moléculas de C3 en C3a y C3b (ver Figura\(\PageIndex{4}\)).
5. Algunas moléculas de C3b se unen a C4b2a, la convertasa C3, para formar C4B2a3b, una convertasa C5 que escinde C5 en C5a y C5b (ver Figura\(\PageIndex{5}\)).
6. C5b se une a la superficie de la célula diana y posteriormente se une a C6, C7, C8 y varios monómeros de C9 para formar C5B6789 n, el Complejo de Ataque a Membrana (MAC) (ver Figura\(\PageIndex{6}\) y Figura\(\PageIndex{7}\)).
Como se mencionó anteriormente, las proteínas de las vías del complemento realizan 6 funciones de defensa innatas beneficiosas. Estos incluyen:
1. Inflamación desencadenante: C5a es la proteína del complemento más potente que desencadene la inflamación. Reacciona con los vasos sanguíneos provocando vasodilatación. También provoca que los mastocitos liberen vasodilatadores como la histamina, aumentando la permeabilidad de los vasos sanguíneos así como aumentando la expresión de moléculas de adhesión en los leucocitos y el endotelio vascular para que los leucocitos puedan exprimirse de los vasos sanguíneos y entrar en el tejido (diapedesis). C5a también hace que los neutrófilos liberen radicales de oxígeno tóxicos para la destrucción extracelular e induce fiebre. En menor medida, C3a y C4a también promueven la inflamación. Como veremos más adelante en esta unidad, la inflamación es un proceso en el que los vasos sanguíneos se dilatan y se vuelven más permeables, permitiendo así que las células de defensa corporal y los químicos de defensa salgan de la sangre y entren en los tejidos.
2. Atraer fagocitos quimiotácticamente al sitio de infección: C5a también funciona como un quimioatrayente para los fagocitos. Los fagocitos se moverán hacia concentraciones crecientes de C5a y posteriormente se unirán, a través de sus receptores CR1, a las moléculas C3b unidas al antígeno. Esto se discutirá con mayor detalle más adelante en esta unidad bajo fagocitosis.
3. Promover la unión de antígenos a fagocitos (mayor unión u opsonización): C3b y en menor medida, C4b puede funcionar como opsoninas, es decir, pueden unir antígenos a fagocitos. Una porción del C3b se une a proteínas y polisacáridos en superficies microbianas; otra porción se une a receptores CR1 en fagocitos, linfocitos B y células dendríticas para mejorar la fagocitosis. (ver Figura\(\PageIndex{8}\)). En la actualidad, la molécula C3b puede unirse a prácticamente cualquier proteína o polisacárido. Las células humanas, sin embargo, producen Factor H que se une a C3b y permite que el Factor I inactive el C3b. Por otro lado, sustancias como LPS en células bacterianas facilitan la unión del Factor B a C3b y esto protege al C3b de la inactivación por el Factor I. De esta manera, C3b no interactúa con nuestras propias células sino que es capaz de interactuar con células microbianas. C3a y C5a aumentan la expresión de receptores C3b en fagocitos y aumentan su actividad metabólica.
4. Causando lisis de bacterias Gram-negativas, células humanas que muestran epítopos extraños y envolturas virales: C5b6789 n, funciona como un Complejo de Ataque de Membrana (MAC). Esto ayuda a destruir bacterias gramnegativas así como células humanas que muestran antígenos extraños (células infectadas por virus, células tumorales, etc.) al provocar su lisis; ver Figura\(\PageIndex{6}\) y Figura\(\PageIndex{7}\). También puede dañar la envoltura de virus envueltos.
5. Sirviendo como segunda señal para activar linfocitos B vívidos durante la inmunidad adaptativa: Algunos C3b se convierten en C3d. C3d se une a los receptores CR2 en los linfocitos B. Esto sirve como una segunda señal para la activación de linfocitos B cuyos receptores de células B acaban de interaccionar con su antígeno correspondiente.
6. Eliminación de complejos inmunes dañinos del cuerpo: C3b y en menor medida, C4b ayudan a eliminar los complejos inmunes dañinos del cuerpo. Los C3b y C4b unen los complejos inmunes a los receptores CR1 en los eritrocitos. Los eritrocitos luego entregan los complejos a macrófagos fijos dentro del bazo y el hígado para su destrucción. Los complejos inmunes pueden conducir a una hipersensibilidad dañina Tipo III, como se discutirá más adelante en la Unidad 5 bajo Hipersensibilidad.
Ejercicio: Preguntas de Pensar-Par-Compartir
- Algunas cápsulas bacterianas son ricas en ácido siálico, un componente común de la glicoproteína de la célula huésped, que tiene afinidad por la proteína del suero H, una proteína reguladora del complemento que conduce a la degradación de C3b.
Describir qué importancia tiene esto en la bacteria resistente a la fagocitosis y por qué.
- S. pyogenes produce una proteasa que escinde la proteína del complemento C5a.
Describir qué importancia tiene esto en la bacteria resistente a la fagocitosis y por qué.
La vía de la lectina
La vía lectina se activa por la interacción de carbohidratos microbianos con lectina de unión a manosa (MBL) o ficolinas que se encuentran en el plasma y los fluidos tisulares. (Las lectinas son proteínas que se unen a carbohidratos). La vía lectina está mediada por dos grupos de proteínas que se encuentran en el plasma de la sangre y en los fluidos tisulares:
1. Lectina de unión a manosa (MBL) - también conocida como proteína de unión a manosa o MBP. MBL es un receptor soluble de reconocimiento de patrones que se une a diversos carbohidratos microbianos como los ricos en manosa o fucosa, y a N-acetilglucosamina (NAG). Estos glicanos son comunes en glicoproteínas microbianas y glicolípidos pero raros en los humanos. La MBL es sintetizada por el hígado y liberada al torrente sanguíneo como parte de la respuesta de fase aguda que se discutirá más adelante en esta unidad. El MBL es equivalente a C1q en la vía clásica del complemento.
Las ficolinas son similares en su estructura a MBL y se unen a carbohidratos microbianos como N-acetilglucosamina (NAG), ácidos lipoteicoicos y lipopolisacáridos (LPS). La ficolina también es equivalente a C1q en la vía clásica del complemento.
2. Tanto la lectina de unión a manosa (MBL) como la ficolina forman complejos con serina proteasas asociadas a MBL llamadas MASP1 y MASP2, que son equivalentes a C1r y C1s de la vía clásica.
a. La unión de la MBL (o la ficolina) al carbohidrato microbiano activa la MASP2 asociada dándole la actividad enzimática para dividir C4 en C4a y C4b (ver Figura\(\PageIndex{9}\) A y Figura\(\PageIndex{9}\) B).
b. C2 luego se une a C4b y se escinde por MASP2 en C2a y C2b (ver Figura\(\PageIndex{10}\) A y Figura\(\PageIndex{10}\) B).
c. C4b y C2a se combinan para formar C4B2a, la convertasa C3. La convertasa C3 ahora puede escindir cientos de moléculas de C3 en C3a y C3b (ver Figura\(\PageIndex{11}\)).
d. Algunas moléculas de C3b se unen a C4b2a, la convertasa C3, para formar C4B2a3b, una convertasa C5 que escinde C5 en C5a y C5b (ver Figura\(\PageIndex{12}\)).
e. C5b se une a la superficie de la célula diana y posteriormente se une a C6, C7, C8 y varios monómeros de C9 para formar C5b6789 n, el Complejo de Ataque a Membrana (MAC) (ver Figura\(\PageIndex{6}\) y Figura\(\PageIndex{7}\)).
Los resultados beneficiosos de las proteínas activadas del complemento son los mismos que en la ruta clásica del complemento anterior. Las proteínas del complemento:
1. Inflamación desencadenante: C5a>C3A>C4a;
2. Atraer quimiotácticamente los fagocitos al sitio de infección: C5a;
3. Promover la unión de antígenos a fagocitos a través de una mayor unión u opsonización: C3b>C4b;
4. Causar lisis de bacterias Gram-negativas y células humanas que presentan epítopos extraños: MAC;
5. Servir como segunda señal para la activación de linfocitos B vívidos): C3d; y
6 Eliminar del cuerpo los complejos inmunes dañinos: C3b>C4b.
La vía alternativa del complemento
La vía alternativa del complemento está mediada por C3b, producida ya sea por la vía clásica o lectina o a partir de hidrólisis C3 por agua. (El agua puede hidrolizar C3 y formar C3i, una molécula que funciona de manera similar a C3b.)
La activación de la vía alternativa del complemento comienza cuando C3b (o C3i) se une a la pared celular y a otros componentes superficiales de los microbios. C3b también puede unirse a anticuerpos IgG. El factor B de proteína de vía alternativa se combina entonces con el C3b unido a células para formar C3bB. El Factor D luego divide el Factor B unido en Bb y Ba, formando C3bbB. Una proteína sérica llamada properdina luego se une al Bb para formar C3bBbp que funciona como una convertasa C3 (ver Figura\(\PageIndex{13}\)) capaz de dividir enzimáticamente cientos de moléculas de C3 en C3a y C3b. Ahora se activa la vía alternativa del complemento.
Algunos de los C3b posteriormente se unen a algunos de los C3bBB para formar C3bB3b, una convertasa C5 capaz de dividir las moléculas de C5 en C5a y C5b (ver Figura\(\PageIndex{14}\)). A partir de aquí, la vía alternativa del complemento es idéntica a las otras vías del complemento.
Los resultados beneficiosos son los mismos que en la ruta clásica del complemento anterior. Las proteínas del complemento:
- Inflamación desencadenante: C5a>C3A>C4a;
- Atraer quimiotácticamente los fagocitos al sitio de infección: C5a;
- Promover la unión de antígenos a fagocitos a través de una mayor unión u opsonización: C3b>C4b;
- Causar lisis de bacterias Gram-negativas, células humanas que muestran epítopos extraños y envolturas virales: MAC; y
- Servir como segunda señal para la activación de linfocitos B vívidos): C3d;
- Eliminar los complejos inmunes dañinos del cuerpo: C3b>C4b.
Tenga en cuenta que en la Unidad 3, aprendimos varios mecanismos que diversas bacterias utilizan para resistir las vías del complemento del cuerpo. Al resistir estas defensas inmunes innatas inmediatas, algunas bacterias tienen más posibilidades de colonizar a su huésped.
Resumen
- Las proteínas del sistema del complemento circulan en forma inactiva, pero en respuesta al reconocimiento de componentes moleculares de microorganismos, se activan secuencialmente, trabajando en una cascada donde en la unión de una proteína promueve la unión de la siguiente proteína en la cascada.
- Existen 3 vías del complemento que conforman el sistema del complemento: la vía clásica del complemento, la vía lectina y la vía alternativa del complemento.
- La vía clásica del complemento se inicia por la activación de C1. C1 se activa principalmente interactuando con la porción Fc de las moléculas de anticuerpo IgG o IgM después de que se hayan unido a su antígeno específico. C1 también es capaz de unirse directamente a las superficies de algunos patógenos así como con la proteína C reactiva (PCR) que se produce durante la respuesta de fase aguda de la inmunidad innata.
- La vía lectina se activa por la interacción de carbohidratos microbianos (lectinas) con lectina de unión a manosa (MBL) o ficolinas que se encuentran en el plasma y fluidos tisulares.
- La vía alternativa del complemento se activa mediante la unión de C3b a superficies microbianas y a moléculas de anticuerpos.
- Todas las vías del complemento realizan las mismas 6 funciones benéficas de defensa innatas.
- Las proteínas del complemento C5a y, en menor medida, C3a y C4a desencadenan vasodilatación e inflamación para entregar células de defensa y químicos de defensa al sitio de la infección.
- La proteína del complemento C5a también funciona como quimioatrayente para los fagocitos.
- Las proteínas del complemento C3b y en menor medida, C4b pueden funcionar como opsoninas, es decir, pueden unir antígenos a fagocitos.
- Las proteínas del complemento C5B6789n, funcionan como un Complejo de Ataque a Membrana (MAC) que causa la lisis de bacterias Gram-negativas, células humanas que muestran epítopos extraños y envolturas virales.
- La proteína del complemento C3d sirve como segunda señal para activar los linfocitos B vívidos durante la inmunidad adaptativa.
- Las proteínas del complemento C3b y en menor medida, C4b ayudan a eliminar los complejos inmunes dañinos del organismo.