
19.4: Replicación de ADN en células procariotas
- Page ID
- 55474

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Describa brevemente el proceso de replicación del ADN.
- Exponer por qué el ADN solo se puede sintetizar en una dirección de 5' a 3'.
- Anotar la función de las siguientes enzimas en la replicación bacteriana del ADN:
- ADN polimerasaIII
- ADN polimerasa II
- ADN helicasa
- primasa
- ADN ligasa
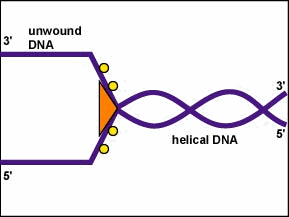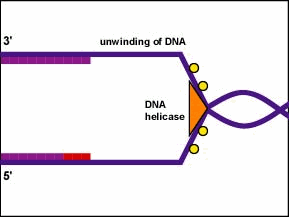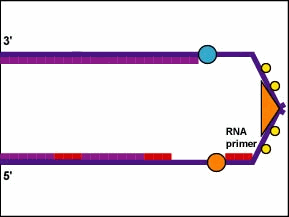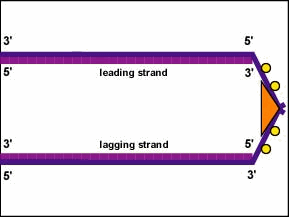
En general, el ADN se replica desenrollando la hélice, separación de cadenas por ruptura de los enlaces de hidrógeno entre las cadenas complementarias y síntesis de dos nuevas cadenas por emparejamiento de bases complementarias. La replicación comienza en un sitio específico en el ADN llamado origen de replicación (ori C). La replicación del ADN es bidireccional desde el origen de la replicación. Para comenzar la replicación del ADN, las enzimas desenrolladas llamadas helicasas de ADN hacen que segmentos cortos de las dos cadenas de ADN parentales se desenrollen y se separen entre sí en el origen de la replicación para formar dos horquillas de replicación en forma de “Y”. Estas horquillas de replicación son el sitio real de la copia del ADN (Figura\(\PageIndex{1}\)).

Todas las proteínas involucradas en la replicación del ADN se agregan en las horquillas de replicación para formar un complejo de replicación llamado replisoma (Figura\(\PageIndex{2}\)).
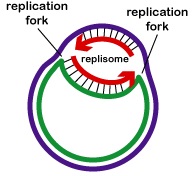
Las proteínas de unión monocatenaria se unen a las regiones monocatenarias para que las dos cadenas no se vuelvan a unir. El desenrollado de la hélice de doble hebra genera superbobinas positivas antes de la horquilla de replicación. Las enzimas llamadas topoisomerasas contrarrestan esto produciendo roturas en el ADN y luego se reunen a ellas para formar superbobinas negativas con el fin de aliviar este estrés en la molécula helicoidal durante la replicación.

A medida que las cadenas continúan desenrollándose y separándose en ambas direcciones alrededor de toda la molécula de ADN, se producen nuevas cadenas complementarias por el enlace de hidrógeno de los nucleótidos de ADN libres con los de cada cadena parental. A medida que los nuevos nucleótidos se alinean frente a cada cadena parental mediante enlaces de hidrógeno, las enzimas llamadas ADN polimerasas se unen a los nucleótidos por medio de enlaces fosfodiéster. En realidad, los nucleótidos que se alinean por emparejamiento de bases complementarias son desoxinucleótidos trifosfatos, compuestos por una base nitrogenada, desoxirribosa y tres fosfatos. A medida que se forma el enlace fosfodiéster entre el grupo fosfato 5' del nuevo nucleótido y el OH 3' del último nucleótido en la cadena de ADN, se eliminan dos de los fosfatos proporcionando energía para la unión (Figura\(\PageIndex{3}\)). Al final, cada hebra parental sirve como molde para sintetizar una copia complementaria de sí misma, dando como resultado la formación de dos moléculas de ADN idénticas (Figura\(\PageIndex{4}\)).
En las bacterias, las proteínas Par funcionan para separar los cromosomas bacterianos a polos opuestos de la célula durante la división celular. Se unen al origen de replicación del ADN y físicamente tiran o empujan los cromosomas separados, similar al aparato mitótico de las células eucariotas. Las proteínas Fts, como FtsK en el divisoma, también ayudan a separar el cromosoma bacteriano replicado.

Animación: Replicación del ADN mediante emparejamiento de bases complementarias. A medida que las cadenas de ADN se desenrollan y se separan, se producen nuevas cadenas complementarias por el enlace de hidrógeno de los nucleótidos de ADN libres con los de cada cadena parental. A medida que los nuevos nucleótidos se alinean frente a cada cadena parental mediante enlaces de hidrógeno, las enzimas llamadas ADN polimerasas se unen a los nucleótidos por medio de enlaces fosfodiéster. Aquí no se muestra la ADN polimerasa responsable de estos eventos.
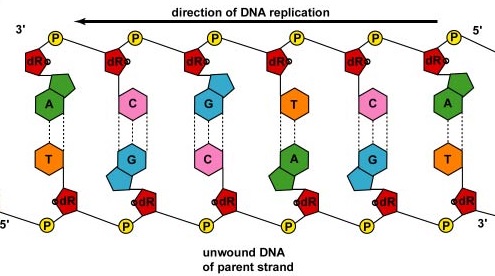
En realidad, la replicación del ADN es más complicada que esto debido a la naturaleza de las ADN polmerasas. Las enzimas ADN polimerasa solo son capaces de unir el grupo fosfato en el carbono 5' de un nuevo nucleótido al grupo hidroxilo (OH) del carbono 3' de un nucleótido que ya está en la cadena. Como resultado, el ADN solo se puede sintetizar en una dirección 5' a 3' mientras se copia una cadena parental que corre en una dirección de 3' a 5'.
Recuerde, como se mencionó anteriormente, cada hebra de ADN tiene dos extremos. El extremo 5' del ADN es el que tiene el grupo fosfato terminal en el carbono 5' de la desoxirribosa; el extremo 3' es el que tiene un grupo hidroxilo terminal (OH) en la desoxirribosa del carbono 3' de la desoxirribosa. Los dos hilos son antiparalelos, es decir, corren en direcciones opuestas. Por lo tanto, una hebra padre, la que corre de 3' a 5' y se llama la hebra principal, se puede copiar directamente a lo largo de toda su longitud (Figura\(\PageIndex{5}\)).

Sin embargo, la otra cadena parental -la que corre 5' a 3' y se llama la hebra rezagada- debe copiarse discontinuamente en fragmentos cortos (fragmentos de Okazaki) de alrededor de 100-1000 nucleótidos cada uno a medida que el ADN se desenrolla. Esto ocurre, como se mencionó anteriormente, en el replisoma. La hebra de ADN retrasada sale de la cadena principal y esto permite que el replisoma se mueva a lo largo de ambas hebras tirando del ADN a medida que ocurre la replicación. Es el ADN real, no la ADN polimerasa que se mueve durante la replicación bacteriana del ADN (Figura\(\PageIndex{2}\)).
Además, las enzimas ADN polimerasa no pueden comenzar una nueva cadena de ADN desde cero. Solo pueden unir nuevos nucleótidos al grupo OH 3' de un nucleótido en una cadena preexistente. Por lo tanto, para iniciar la síntesis de la cadena principal y cada fragmento de ADN de la hebra rezagada, se requiere un complejo de ARN polimerasa llamado primasa. La primasa, que es capaz de unir nucleótidos de ARN sin requerir una cadena preexistente de ácido nucleico, primero agrega varios nucleótidos de ARN complementarios opuestos a los nucleótidos de ADN en la cadena parental. Esto forma lo que se llama un cebador de ARN (Figura\(\PageIndex{6}\)).

Luego, la ADN polimerasa III reemplaza a la primasa y es capaz de agregar nucleótidos de ADN al cebador de ARN (Figura\(\PageIndex{7}\)).
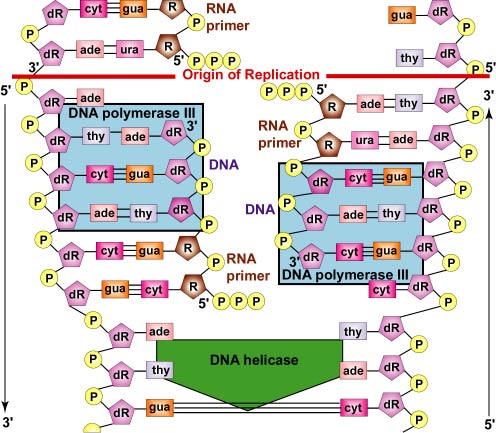
Posteriormente, la ADN polimerasa II digiere el cebador de ARN y reemplaza los nucleótidos de ARN del cebador con los nucleótidos de ADN adecuados para llenar el hueco (Figura\(\PageIndex{8}\)).

Animación: Replicación de hebras de ADN líderes y rezagadas. La cadena principal se realiza continuamente en una dirección 5' a 3' por la ADN polimerasa III a medida que la helicasa de ADN desenrolla la hélice de ADN parental. Sin embargo, debido a que las cadenas de ADN parentales son antiparalelas, la hebra rezagada debe hacerse en fragmentos cortos. La ARN polimerasa (primasa) sintetiza un cebador de ARN corto el cual es extendido por la ADN polimerasa III. A continuación, la ADN polimerasa II digiere el cebador de ARN y lo reemplaza con ADN. Finalmente, la ADN ligasa une los fragmentos de la hebra rezagada.
Existe una gran cantidad de información genética en el cromosoma bacteriano. Por ejemplo Escherichia coli, la bacteria más estudiada de todas, tiene un genoma que contiene 4,639.221 pares de bases, que codifican al menos 4288 proteínas.
Resumen
- Durante la replicación del ADN, cada cadena parental actúa como molde para la síntesis de la otra cadena por medio de apareamiento de bases complementarias.
- El apareamiento de bases complementario se refiere a nucleótidos de ADN con la base adenina formando únicamente enlaces de hidrógeno con nucleótidos que tienen la base timina (A-T). Asimismo, los nucleótidos con la base guanina pueden unirse por hidrógeno solo con nucleótidos que tienen la base citosina (G-C).
- Cada cadena de ADN tiene dos extremos. El extremo 5' del ADN es el que tiene el grupo fosfato terminal en el carbono 5' de la desoxirribosa; el extremo 3' es el que tiene un grupo hidroxilo terminal (OH) en la desoxirribosa del carbono 3' de la desoxirribosa.
- Para sintetizar las dos cadenas de desoxirribonucleótidos durante la replicación del ADN, las enzimas ADN polimerasa involucradas solo son capaces de unir el grupo fosfato en el carbono 5' de un nuevo nucleótido al grupo hidroxilo (OH) del carbono 3' de un nucleótido ya en la cadena.
- Si bien las dos cadenas de ADN son complementarias, están orientadas en direcciones opuestas entre sí. Se dice que una cadena corre 5' a 3'; la cadena de ADN opuesta corre antiparalela, o 3' a 5'.
- Para comenzar la replicación del ADN, las enzimas desenrolladas llamadas helicasas de ADN hacen que segmentos cortos de las dos cadenas de ADN parentales se desenrollen y se separen entre sí en el origen de la replicación para formar dos horquillas de replicación en forma de “Y”.
- Las proteínas de unión monocatenaria se unen a las regiones monocatenarias ahora desapareadas para que las dos cadenas no se vuelvan a unir.
- A medida que las cadenas continúan desenrollándose y separándose en ambas direcciones alrededor de toda la molécula de ADN, se producen nuevas cadenas complementarias por el enlace de hidrógeno de los nucleótidos de ADN libres con los de cada cadena parental.
- A medida que los nuevos nucleótidos se alinean frente a cada cadena parental mediante enlaces de hidrógeno, las enzimas llamadas ADN polimerasas se unen a los nucleótidos por medio de enlaces fosfodiéster.
- Los dos hilos son antiparalelos, es decir, corren en direcciones opuestas. Por lo tanto, una hebra padre, la que corre de 3' a 5' y se llama la hebra principal, puede copiarse directamente en toda su longitud. Sin embargo, la otra cadena parental -la que corre 5' a 3' y se llama la cadena rezagada- debe copiarse discontinuamente en fragmentos cortos (fragmentos de Okazaki) de alrededor de 100-1000 nucleótidos cada uno a medida que el ADN se desenrolla.
- Además, las enzimas ADN polimerasa no pueden comenzar una nueva cadena de ADN desde cero. Solo pueden unir nuevos nucleótidos al grupo OH 3' de un nucleótido en una cadena preexistente. Por lo tanto, para iniciar la síntesis de la cadena principal y cada fragmento de ADN de la hebra rezagada, se requiere un complejo de ARN polimerasa llamado primasa. La primasa, que es capaz de unir nucleótidos de ARN sin requerir una cadena preexistente de ácido nucleico, primero agrega varios nucleótidos de ARN complementarios opuestos a los nucleótidos de ADN en la cadena parental formando lo que se llama cebador de ARN.
- Luego, la ADN polimerasa III reemplaza a la primasa y es capaz de agregar nucleótidos de ADN al cebador de ARN. Posteriormente, la ADN polimerasa II digiere el cebador de ARN y reemplaza los nucleótidos de ARN del cebador con los nucleótidos de ADN adecuados para llenar el vacío.
- Los propios fragmentos de ADN son enganchados por la enzima ADN ligasa para completar el proceso.