7.7A: Procesamiento de ARNm
- Page ID
- 60760

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
El pre-ARNm eucariota recibe una caperuza 5' y una cola poli (A) en 3" antes de que se eliminen los intrones y se considere que el ARNm está listo para la traducción.
OBJETIVOS DE APRENDIZAJE
Esbozar los pasos del procesamiento previo al ARNm
Claves para llevar
Puntos Clave
- Se agrega una tapa de 7-metilguanosina al extremo 5' del pre-ARNm mientras la elongación aún está en progreso. La caperuza 5' protege el ARNm naciente de la degradación y ayuda en la unión al ribosoma durante la traducción.
- Una cola de poli (A) se agrega al extremo 3" del pre-ARNm una vez que se completa la elongación. La cola poli (A) protege al ARNm de la degradación, ayuda en la exportación del ARNm maduro al citoplasma y participa en la unión de proteínas involucradas en el inicio de la traducción.
- Los intrones se eliminan del pre-ARNm antes de que el ARNm se exporte al citoplasma.
Términos Clave
- intrón: una porción de un gen dividido que se incluye en transcritos pre-ARN pero que se elimina durante el procesamiento del ARN y se degrada rápidamente
- resto: un segmento específico de una molécula
- spliceosoma: un complejo dinámico de subunidades de ARN y proteínas que elimina intrones del ARNm precursor
Procesamiento de pre-ARNm
El pre-ARNm eucariota se somete a un procesamiento extenso antes de que esté listo para ser traducido. Los pasos adicionales involucrados en la maduración del ARNm eucariota crean una molécula con una vida media mucho más larga que un ARNm procariota. Los ARNm eucariotas duran varias horas, mientras que el ARNm típico de E. coli no dura más de cinco segundos.
Los pre-ARNm se recubren primero en proteínas estabilizadoras de ARN; estas protegen al pre-ARNm de la degradación mientras se procesa y exporta fuera del núcleo. Los tres pasos más importantes del procesamiento del pre-ARNm son la adición de factores estabilizantes y de señalización en los extremos 5' y 3' de la molécula, y la eliminación de secuencias intervinientes que no especifican los aminoácidos apropiados. En raras ocasiones, la transcripción del ARNm puede ser “editada” después de ser transcrita.
Tapado de 5′
Mientras el pre-ARNm aún se está sintetizando, se agrega una tapa de 7-metilguanosina al extremo 5' del transcrito en crecimiento mediante un enlace fosfato de 5' a 5'. Este resto protege al ARNm naciente de la degradación. Además, los factores de iniciación involucrados en la síntesis de proteínas reconocen la tapa para ayudar a iniciar la traducción por los ribosomas.
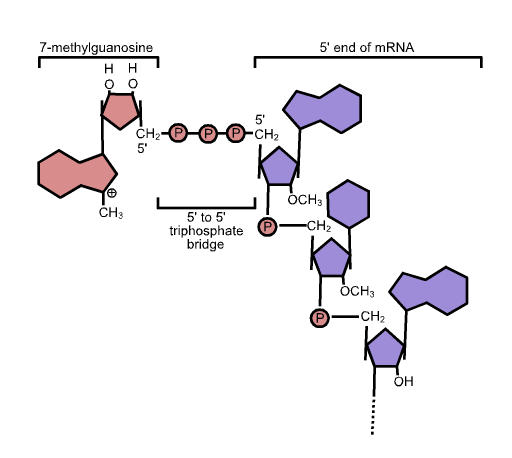
Cola Poly-A de 3′
Mientras que la ARN polimerasa II todavía se está transcribiendo aguas abajo del extremo apropiado de un gen, el pre-ARNm es escindido por un complejo proteico que contiene endonucleasa entre una secuencia consenso de AAUAAA y una secuencia rica en Gu. Esto libera el pre-ARNm funcional del resto del transcrito, que aún está unido a la ARN Polimerasa. Una enzima llamada poli (A) polimerasa (PAP) es parte del mismo complejo proteico que escinde el pre-ARNm e inmediatamente agrega una cadena de aproximadamente 200 nucleótidos A, llamada cola poli (A), al extremo 3' del pre-ARNm recién escindido. La cola poli (A) protege al ARNm de la degradación, ayuda en la exportación del ARNm maduro al citoplasma y participa en la unión de proteínas involucradas en el inicio de la traducción.

La Poli (A) Polimerasa agrega una cola 3′ poli (A) al pre-ARNm. : El pre-ARNm se escinde del resto del transcrito en crecimiento antes de que la ARN Polimerasa II haya dejado de transcribir. Esta escisión se realiza mediante un complejo proteico que contiene endonucleasa que se une a una secuencia de AAUAAA aguas arriba del sitio de escisión y a una secuencia rica en Gu aguas abajo del sitio de corte. Inmediatamente después de la escisión, la Poli (A) Polimerasa (PAP), que también forma parte del complejo proteico, cataliza la adición de hasta 200 nucleótidos A al extremo 3' del pre-ARNm recién escindido.
Empalme pre-mRNA
Los genes eucariotas están compuestos por exones, que corresponden a secuencias codificadoras de proteínas (ex -on significa que son ex presionadas), y secuencias intervinientes llamadas intrones (int -ron denota su papel int ervening), que pueden estar involucradas en la regulación génica, pero se eliminan del pre-ARNm durante el procesamiento. Las secuencias de intrones en ARNm no codifican proteínas funcionales.
Descubrimiento de intrones
El descubrimiento de intrones fue una sorpresa para los investigadores en la década de 1970 que esperaban que los pre-ARNm especificaran secuencias de proteínas sin procesamiento adicional, como habían observado en procariotas. Los genes de los eucariotas superiores contienen muy a menudo uno o más intrones. Si bien estas regiones pueden corresponder a secuencias reguladoras, la importancia biológica de tener muchos intrones o tener intrones muy largos en un gen no está clara. Es posible que los intrones ralenticen la expresión génica porque lleva más tiempo transcribir pre-ARNm con muchos intrones. Alternativamente, los intrones pueden ser restos de secuencia no funcionales sobrantes de la fusión de genes antiguos a lo largo de la evolución. Esto está respaldado por el hecho de que los exones separados a menudo codifican subunidades o dominios de proteínas separadas. En su mayor parte, las secuencias de intrones pueden mutarse sin afectar en última instancia al producto proteico.
Procesamiento de intrones
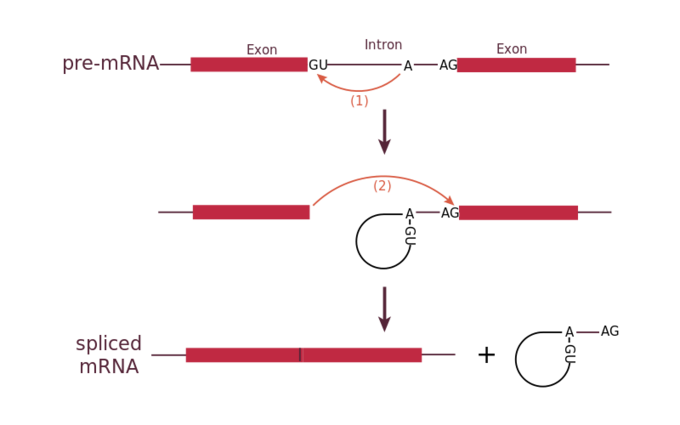
Todos los intrones en un pre-ARNm deben eliminarse completa y precisamente antes de la síntesis de proteínas. Si el proceso falla incluso por un solo nucleótido, el marco de lectura de los exones reunidos se desplazaría y la proteína resultante sería disfuncional. El proceso de eliminación de intrones y reconexión de exones se llama empalme. Los intrones se eliminan y degradan mientras el pre-ARNm aún se encuentra en el núcleo. El empalme se produce mediante un mecanismo específico de secuencia que asegura que los intrones se eliminen y los exones se vuelvan a unir con la precisión y precisión de un solo nucleótido. El corte y empalme de pre-ARNm es conducido por complejos de proteínas y moléculas de ARN llamados spliceosomas.
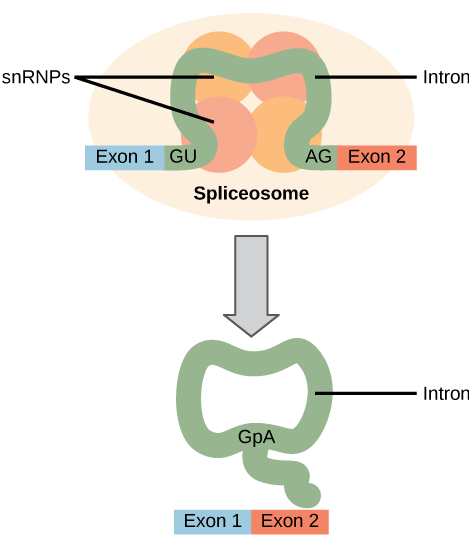
Cada spliceosoma está compuesto por cinco subunidades llamadas snRNP (para ribonucleopartículas nucleares pequeñas, y pronunciadas “snurps”). Cada snRNP es en sí mismo un complejo de proteínas y un tipo especial de ARN que se encuentra solo en el núcleo llamado ARNsn (ARN nucleares pequeños). Los spliceosomas reconocen secuencias en el extremo 5' del intrón porque los intrones siempre comienzan con los nucleótidos GU y reconocen secuencias en el extremo 3' del intrón porque siempre terminan con los nucleótidos AG. El spliceosoma escinde la cadena principal de fosfato de azúcar del pre-ARNm en la G que inicia el intrón y luego une covalentemente esa G a un nucleótido A interno dentro del intrón. Entonces el spliceosme conecta el extremo 3′ del primer exón al extremo 5' del siguiente exón, escindiendo el extremo 3′ del intrón en el proceso. Esto da como resultado el empalme de los dos exones y la liberación del intrón en forma de lariat.