
4.8: Nuestro pasado antiguo- Los primeros homínidos
- Page ID
- 149326

Al final de esta sección, podrás:
Caminando en Dos Pies
El término homínido se refiere a todas las especies consideradas en linaje directo con los humanos, que incluyen los géneros Homo, Australopithecus, Paranthropus y Ardipithecus. Homínidos se refiere a todos los grandes simios modernos y extintos, que incluyen humanos, gorilas, chimpancés y orangutanes y sus antepasados. Se ha entendido que estos términos representan cosas diferentes a lo largo de los años, pero las definiciones que aquí se proporcionan son las más actuales. Si bien todos los homínidos pueden diferir en diferentes formas entre sí, todos comparten un complejo anatómico conductual: la locomoción bípeda.
Los científicos pueden plantear hipótesis sobre cómo se movió una criatura analizando varios aspectos de su morfología. Los braquiadores, animales que se mueven balanceándose de rama en rama, generalmente tienen brazos largos, mientras que los saltadores, animales que impulsan sus cuerpos a través de la fuerza de sus miembros inferiores, tienen patas relativamente largas. Los primates arbóreos tienen brazos y piernas de igual longitud. En la locomoción bípeda, una pierna se llama pierna de postura, y la otra se llama pierna escalonada. Mientras que la pierna de postura está en el suelo, la pierna de paso está fuera del suelo y dando un paso adelante. Durante la marcha normal, ambos pies están en el suelo solo alrededor del 25 por ciento del tiempo. A medida que aumenta la velocidad de locomoción, disminuye el porcentaje de tiempo que ambos pies están en el suelo. Como resultado, durante la mayor parte del tiempo que los organismos bípedos se mueven, su cuerpo se equilibra en una sola de sus piernas (la pierna de postura). Para garantizar que los organismos bípedos no se caigan mientras están equilibrados en su pierna de postura, han sufrido muchos cambios anatómicos desde los primeros antepasados de los homínidos.
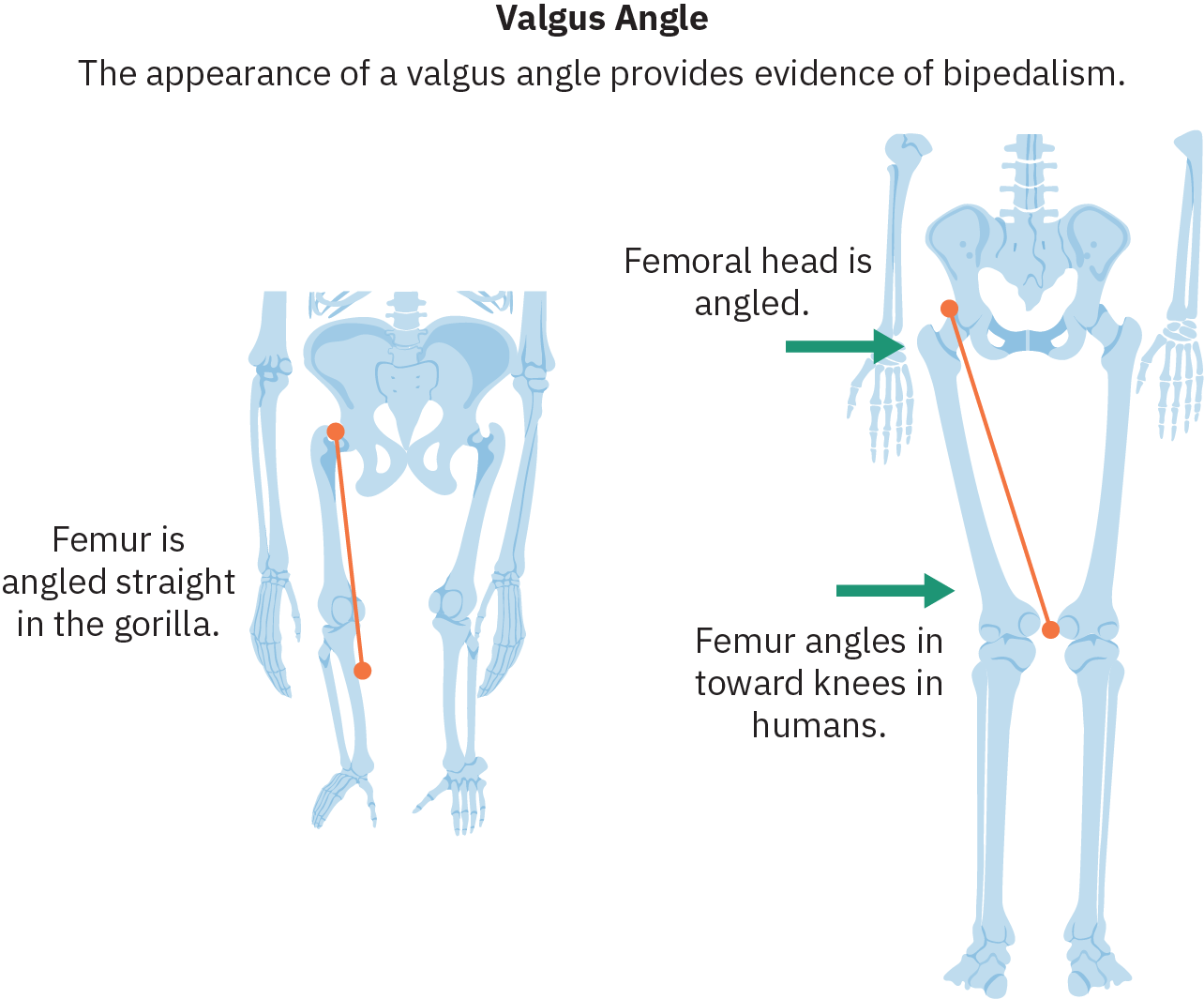
Uno de los cambios anatómicos más importantes que facilitan el bipedalismo exitoso es la inclinación del fémur (hueso de la parte superior de la pierna) hacia adentro en lo que se conoce como ángulo valgo, que posiciona las rodillas y los pies debajo del centro de la pelvis. Los homínidos bípedos también han evolucionado curvas espinales que permiten que las caderas equilibren el peso de la parte superior del cuerpo. La evolución del arco en el pie así como el realineamiento del dedo gordo del pie para que sea paralelo a los otros dedos también es instrumental en la transmisión de peso durante la fase escalonada de la locomoción bípeda.
La evidencia más importante del bipedalismo temprano de los homínidos es proporcionada por el trabajo de la paleoantropóloga inglesa Mary Leakey. En la década de 1980, Mary Leakey descubrió un rastro de 75 pies de huellas hechas por tres individuos bípedos que habían cruzado una gruesa cama de ceniza volcánica húmeda alrededor de 3.5 MYA. Estas huellas fueron encontradas en África Oriental en el sitio de Laetoli. Con base en la fecha y la ubicación, es probable que estas huellas hayan sido realizadas por Australopithecus afarensis. El análisis de las huellas de Laetoli indica una marcha moderna.

La evolución del bipedalismo homínico requirió una compleja reorganización anatómica. Para que la selección natural produzca una cantidad tan tremenda de cambio, los beneficios de estos cambios deben haber sido grandes. Ha habido docenas de hipótesis para estos cambios, que van desde liberar las manos para llevar herramientas, alimentos o descendencia hasta aumentar la eficiencia energética o la termorregulación (la capacidad de mantener la temperatura del cuerpo) al exponer más de la superficie del cuerpo. Ninguna de las hipótesis es comprobable, lo que hace que sea realmente desafiante entender por qué los antepasados de la humanidad hicieron un cambio de comportamiento tan enorme. Las siguientes secciones exploran algunos de los descubrimientos clave de los primeros fósiles de homínidos en los que los antropólogos ven algunos de los primeros indicios de la adaptación del bipedalismo en la historia humana.
Homínidos del Mioceno
Los primeros fósiles de homínidos aparecen en el Mioceno tardío, de 10 a 5 MYA. En algún momento entre 7 MYA y 4 MYA, los homínidos salieron de los árboles y comenzaron a adaptarse más plenamente a un nicho de vida terrestre. Desafortunadamente, la evidencia fósil de este período de tiempo es extremadamente escasa, pero se siguen descubriendo nuevos hallazgos.
Un cráneo completo de Sahelanthropus tchadensis fue encontrado en 2002 por el paleoantropólogo francés Michel Brunet y su equipo en Chad en África Occidental. Sahelanthropus es un simio fósil que vivió aproximadamente 7 MYA y algunos investigadores afirman ser el último ancestro común de humanos y chimpancés. Los estudios genéticos indican que los humanos y los chimpancés divergieron entre sí en algún momento entre 5 MYA y 7 MYA, por lo que esta especie vivió justo en el momento de la divergencia. La capacidad craneal es de apenas 350 centímetros cúbicos (cc), lo que equivale a un chimpancé; la capacidad craneal humana moderna es de aproximadamente 1,400 cc. Sahelanthropus también tiene una cresta de cejas muy grande (el hueso grande sobre los ojos), y la ubicación del foramen magnum, la abertura en la base del cráneo donde la columna vertebral ingresa al cráneo, sugiere que su cabeza no estaba sujeta sobre su columna vertebral y por lo tanto no era bípeda.
Orrorin tugenensis fue encontrado en Kenia en 2001 por el geólogo Martin Pickford del Collège de France y la paleontóloga Brigitte Senut del Museo Nacional de Historia Natural de Francia. Orrorin tugenensis data de aproximadamente 6 MYA. Se propuso que Orrorin fuera un homínido debido a rasgos anatómicos que sugieren bipedalismo. Por ejemplo, la cabeza femoral (la bola grande y redondeada en la parte superior del hueso de la pierna que conecta la pierna con la cadera) es mucho más grande que en los simios cuadrupedos, lo que sugiere que el fémur se estaba utilizando para soportar el peso de la parte superior del cuerpo. Los músculos adheridos al fémur también sugieren movimiento bípedo. Otra característica que sugiere que Orrorin es verdaderamente un homínido son los dientes, que exhiben esmalte dental grueso y pequeños molares cuadrados, al igual que los humanos modernos.
Homínidos del Plioceno
La época del Plioceno se extendió de 5 MYA a 1.8 MYA. Los fósiles del Plioceno muestran evidencia de la evolución de los homínidos claramente bípedos. También muestran evidencia de comportamiento cultural claro, aunque primitivo. Climáticamente, el Plioceno era más frío que el Mioceno anterior, lo que resultó en cambios en el nivel del mar y un aumento del hielo en los polos, abriendo algunas áreas previamente inaccesibles. Durante este período, América del Norte y del Sur se conectaron a través del Istmo de Panamá, y un puente terrestre a través del Estrecho de Bering apareció entre Alaska y Siberia.
Ardipithecus ramidus
Ardipithecus ramidus fue encontrado en Etiopía en 1992 por el paleoantropólogo estadounidense Tim White y data de aproximadamente 4.4 MYA. Esta es la primera especie de homínidos descubiertos que data de la época del Plioceno. Con base en la posición delantera del foramen magnum, se puede concluir que Ardipithecus fue bípedo. Además, los huesos de la parte superior del brazo son muy pequeños, lo que sugiere que los brazos no se utilizaron para soportar el peso durante la locomoción. Ardipithecus posee numerosos rasgos, como esmalte dental delgado, evidencia de un canino reducido y un dedo gordo oponible. Derivado de este último rasgo, muchos creen que Ardipithecus era bípedo en el suelo y cuadrupédico en los árboles. Esta hipótesis se sustenta en el hecho de que los huesos fósiles se encontraron en ambientes relativamente densamente boscosos. El canino reducido es un rasgo derivado que aparece incluso antes que A. ramidus y no es lo que normalmente veríamos en machos de simios africanos que tienen grandes caninos intimidantes. Las hipótesis actuales sugieren que con el tiempo los caninos más pequeños se volvieron dominantes cuando hubo menos necesidad de mostrar agresión junto con una preferencia femenina por machos con temperamentos más suaves (Suwa, G., et al. 2021).

Los Australopitecinos Robustos y Graciles
En las siguientes secciones se examinarán diversas especies australopitecinas que tenían diversas características físicas relacionadas con la morfología de los dientes y el cráneo. Con base en estas características, los paleoantropólogos clasificaron estas especies en formas graciles y robustas, como se ilustra en la Figura 4.33. La especie Gracile tuvo una proyección más pronunciada de la mandíbula (prognatismo), mejillas menos acampanadas sin cresta sagital y dientes y mandíbulas más pequeñas. La cresta sagital en los robustos australopitecinos acomodó grandes músculos de la mandíbula temporal para masticar materiales vegetales duros.
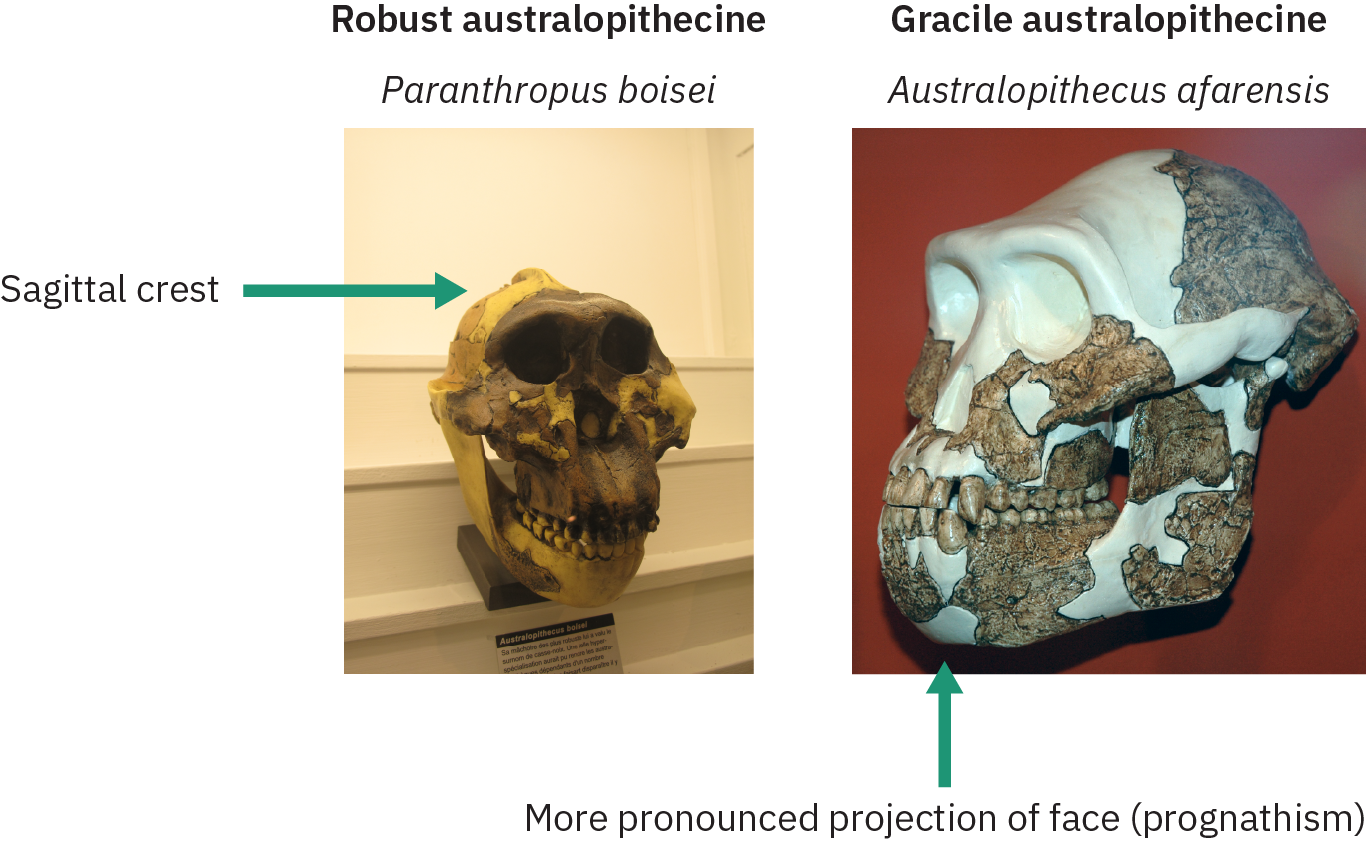
Las especies consideradas como graciles incluyen Australopithecus anamensis, A. afarensis, A. africanus, A. garhi, y A. sediba. Los australopitecinos robustos (clasificados bajo el género Paranthropus) incluyen Paranthropus robustus, P. boisei y P. aethiopicus. La especie gracile emergió alrededor de 4 MYA y desapareció 2 MYA, mientras que las especies robustas continuaron existiendo por otro millón de años. En las siguientes secciones se analizarán primero algunas de las formas graciles del australopitecino, seguidas de las formas robustas.
Australopithecus africanus
Australopithecus africanus fue el primer australopitecino descubierto, en 1924, y fue descrito por el anatomista y antropólogo australiano Raymond Dart, quien encontró el fósil en una caja de fósiles que le enviaron trabajadores de canteras de cal en un sitio llamado Taung en Sudáfrica. El ejemplar más notable de la caja fue una calavera de un niño, que Dart tuvo que astillar de la piedra en la que estaba incrustada. Dart tardó cuatro años en separar los dientes. El cráneo ahora se conoce como el cráneo de Taung o niño Taung. Dart argumentó que el niño Taung representa “una raza extinta de simios intermedia entre los antropoides vivos y el hombre” (Wayman 2011). Señaló que el cráneo era largo y estrecho, no redondeado como en los humanos modernos, y su cerebro promedió apenas 422 cc, equivalente a un chimpancé. Sin embargo, el niño Taung no poseía crestas de cejas, tenía órbitas circulares, y tenía mínimo prognatismo, así como pequeños caninos y sin diastema (espacio en la mandíbula para que los caninos grandes se colocaran cuando se cierra la boca). Estos últimos rasgos son todos análogos a los humanos modernos. Lo más importante, Dart señaló que la posición delantera del foramen magnum indicó que el cráneo estaba equilibrado en la parte superior de la columna vertebral, sugiriendo bipedalismo y una postura erguida.

Australopithecus afarensis
En 1973, una buena porción de un esqueleto (alrededor del 40 por ciento) fue encontrada en la región de Afar de Etiopía por el paleoantropólogo estadounidense Donald Johanson. Llamó al esqueleto Lucy, después de una canción de los Beatles. Se fechó alrededor de 3.75—2.8 MYA y se determinó que era miembro de la especie Australopithecus afarensis. Como todos los fósiles descubiertos recientemente, Lucy recibió un número de identificación o acceso, KNM-AL-288. El acrónimo KNM significa el Museo Nacional de Kenia, donde se encuentra el fósil, y AL significa la localidad Afar donde se encontró el fósil. Desde entonces, se han encontrado más ejemplares de esta especie en Kenia, Tanzania y Etiopía, todos en África Oriental.

Australopithecus afarensis data de 3.9 a 2.9 MYA con una capacidad endocranial de alrededor de 400 cc, que es aproximadamente la misma que un chimpancé común. Hay dos características morfológicas que proporcionan evidencia de que A. afarenis se movía más como un gran simio que como un humano. Primero, tenía brazos sustancialmente más largos que los de los humanos modernos. Los brazos largos generalmente se encuentran en animales que cuelgan de ramas, lo que sugiere que A. afarensis también exhibió este comportamiento. Además, A. afarensis posee huesos de dedos y pies largos y curvos, otra característica de los animales que cuelgan de las ramas. Sin embargo, hay una característica morfológica importante de A. afarensis que sugiere que esta especie puede haberse movido algo como los humanos modernos. La forma de la pelvis de A. afarensis (huesos de la cadera) se parece sustancialmente más a la de un humano moderno que a un simio; en lugar de que los huesos de la cadera sean largos y estrechos, son cortos y anchos. La mayoría de los paleoantropólogos creen que este cambio en la forma pélvica indica que A. afarensis se movió como lo hacen los humanos modernos, en dos piernas. Si bien A. afarensis puede haberse locomocionado bipedalmente, las diferencias morfológicas entre A. afarensis y los humanos modernos sugieren que no se movieron exactamente de la misma manera. El consenso actual es que A. afarensis fue tanto morada en árboles como bípeda. Otra evidencia anatómica de bipedalismo incluye una posición más anterior del foramen magno y el ángulo de la cabeza y cuello femorales.
Australopithecus garhi
También se encuentra en Etiopía, Australopithecus garhi data de aproximadamente 2.5 MYA. Su capacidad craneal es ligeramente mayor que A. afarensis, a 450 cc. Australopithecus garhi tiene incisivos que son más grandes que los de cualquiera de los australopitecinos conocidos o Homo. Aún no se conoce la función de los incisivos grandes. El aspecto más emocionante de A. garhi es que proporciona evidencia del primer uso de herramientas de piedra por un homínido. Específicamente, se encontraron fósiles de A. garhi con huesos fósiles de rumiantes, como antílopes, que mostraban numerosas marcas de corte. Las marcas de corte se hacen en los huesos mediante el proceso de quitar la carne de los huesos con herramientas de piedra o metal. Con base en este hallazgo, los antropólogos biológicos han planteado la hipótesis de que A. garhi utilizó algún tipo de herramienta de piedra para la carnicería.
Australopithecus sediba
En 2008, el hueso de clavícula de Australopithecus sediba fue descubierto por Matthew Berger, el hijo de nueve años del paleontólogo estadounidense Lee Berger, en Malapa, Sudáfrica. Una excavación adicional en una característica de cueva descubrió dos esqueletos parciales, uno de una hembra adulta y el otro un juvenil más joven. A. sediba es considerada una especie importante porque aparece en el registro fósil alrededor de la época de la primera emergencia del género Homo alrededor de 2 mya. La clasificación de A. sediba fue inicialmente difícil de determinar, debido a sus complejas características superpuestas, que incluyen columna vertebral, pelvis, manos y dientes similares a humanos y un pie similar a un chimpancé. Esta combinación de rasgos sugiere tanto la escalada de árboles como las adaptaciones bípedas. Después de estudiar las características colectivamente, los antropólogos clasificaron a A. sediba como una especie de Australopithecus. Se considera un antepasado directo de Homo erectus y Homo ergaster, los cuales se discuten en el Capítulo 5, El género Homo y la emergencia de nosotros. Se cree que A. sediba podría ser descendiente de A. africanus, lo que sugiere que la especie puede ser un callejón sin salida dentro del linaje para los humanos. Su clasificación y relación con el género Homo probablemente seguirá siendo muy debatida.
Paranthropus robustus
Trece años después del descubrimiento de Raymond Dart, el paleontólogo y médico sudafricano Robert Broom descubrió Paranthropus robustus en un sitio llamado Kromdraai en Sudáfrica. La diferencia más obvia entre los respectivos fósiles de Dart y Broom, A. africanus y P. robustus, es que la morfología del fósil de Broom es mucho mayor. Sus características incluyen una cresta sagital y un arco cigomático acampanado para la fijación de un gran músculo temporal para masticar una dieta que depende de frutos secos y semillas duras. Esta interpretación fue apoyada además por la microscopía electrónica de barrido (SEM), la cual se utilizó para evaluar las marcas grabadas en los dientes. A medida que los dientes aumentaban de tamaño los incisivos y caninos se encogieron, dando a Paranthropus una cara más plana con menos proyección de la mandíbula. Hay quienes argumentan que dependiendo del ambiente y la localidad, algunos Paranthropus pueden haber sido omnívoros, con dietas variadas similares a las de H. ergaster. (Lee-Thorp, Thackeray y van der Merwe 2000).
Paranthropus boisei
Siguiendo los pasos de Broom, otros científicos comenzaron a buscar fósiles en África Oriental. A partir de 1931, el paleoantropólogo keniano y británico Louis Leakey y su esposa, Mary Leakey, trabajaron en lo que se conoce como el Valle del Rift Oriental, que es un valle de 1.200 millas que se extiende a través de Etiopía, Kenia y Tanzania. Buscaron durante casi 30 años antes de encontrar su primer fósil de homínidos, Paranthropus boisei (OH-5) —originalmente clasificado como Zinjanthropus boisei — en 1959. A menudo se le conoce como el homínido hiperrobusto debido a su mohawk de hueso en la parte superior del cráneo. Otras características incluyen una frente baja o ausente, una cara plana, mandíbulas grandes y grandes sitios de fijación en todo el cráneo para masticar músculos.
Paranthropus aethiopicus
Tenemos poco conocimiento sobre Paranthropus aethiopicus (que se muestra en la Figura 4.37), que ha sido fechado en aproximadamente 2.5 MYA y se conoce como el “cráneo negro”. Se cree que esta especie cae en algún lugar entre los australopitecinos robustos y graciles, teniendo características de ambos. La especie fue descubierta en Etiopía en 1967 por un equipo de expedición francés encabezado por Camille Arambourg e Yves Coppens.

Hitos y Preguntas
Si bien los fósiles descubiertos hasta este momento han proporcionado una pequeña ventana a la historia del pasado de la humanidad, también han planteado simultáneamente numerosos interrogantes. Las preguntas relacionadas con las relaciones filogenéticas y los puntos de divergencia son desafíos para los paleoantropólogos, quienes solo tienen evidencia fósil fragmentaria para construir hipótesis alrededor. Sin embargo, los descubrimientos que se han realizado representan importantes hitos en la comprensión de los antropólogos, aportando pistas que conducirán a los próximos pasos en el viaje humano.
Mini-Actividad de Trabajo de Campo
Levantamiento Peatonal
Realiza una encuesta de peatones para tratar de localizar fósiles cerca de donde vives (trilobites en Nueva York, amonitas en Texas, dientes de tiburón cerca de lechos de ríos, puntas de flecha). Piensa en dónde probablemente encontrarías un fósil y por qué. Trate de extraer uno sin destruir el entorno que lo rodea, lo que proporciona un contexto importante. Intenta averiguar qué tipo de fósil es haciendo algunas investigaciones en Internet. ¿Por qué crees que se conservó este fósil? ¿Qué información facilitaría la búsqueda de fósiles?