
6: Evolución
- Page ID
- 82554

Objetivos de aprendizaje
Después de completar este capítulo, podrás
- Explicar las diferencias en las condiciones ambientales antes y después de la génesis natural de la vida.
- Discutir las diferencias entre creacionismo y evolución como explicación del origen de la vida y las especies.
- Describir la teoría de la evolución por selección natural.
- Explicar el papel de la genética en la comprensión de la evolución y la biodiversidad.
En el Principio...
Con base en datos geológicos y astronómicos, se cree que la Tierra se originó por la condensación del polvo interestelar hace unos 4.5 mil millones de años. Los entornos previos a la vida del planeta eran muy diferentes de lo que existe hoy en día. La atmósfera inicial probablemente resultó de la desgasificación volcánica y su química estuvo dominada por sulfuro de hidrógeno (H 2 S), metano (CH 4), amoníaco (NH 3), dióxido de carbono (CO 2) y otros gases que hoy existen solo en concentraciones traza. En contraste, la atmósfera moderna tiene grandes concentraciones de oxígeno (O 2) y nitrógeno (N 2).
Una de las razones de un cambio profundo en la química atmosférica fue la evolución de los organismos fotosintéticos, que liberan O 2 como producto de desecho de su metabolismo autotrófico. A medida que aumentaba la concentración de O 2, la atmósfera cambió de un ambiente que favorecía las reacciones reductoras (en el que los productos de reacción tienen una ganancia neta en electrones) a uno en el que predominaban las reacciones oxidantes. El sulfuro de hidrógeno, el metano y el amoníaco son compuestos reducidos, pero en una atmósfera rica en O 2, se oxidan a sulfato (SO 4 2—), dióxido de carbono y nitrato (NO3—), respectivamente. Además, el O2 puede participar en reacciones fotoquímicas que producen pequeñas cantidades de ozono (O3). Cuando está presente en la atmósfera superior, el ozono absorbe la radiación ultravioleta solar y con ello protege a los organismos de muchos de los efectos dañinos de este tipo de energía electromagnética.
Se cree que la génesis de la vida en la Tierra ocurrió en un ambiente acuático primordial hace al menos 3.5 mil millones de años. No se sabe exactamente cómo comenzó la vida por primera vez a partir de la materia inanimada, aunque muchos biólogos creen que el proceso fue una ocurrencia espontánea. Es decir, el origen de la vida ocurrió de manera natural, como consecuencia de la existencia de condiciones apropiadas de química, temperatura, presión, energía, y otros factores ambientales.
Como tal, el origen de la vida podría haber ocurrido como una serie de eventos aleatorios ocurridos en condiciones adecuadas. Algunos biólogos, sin embargo, creen que la génesis podría haber tenido lugar de una manera más intencional, bajo la influencia de reacciones autocatalíticas (autocatalizadas) que favorecieron la síntesis y persistencia de sustancias químicas orgánicas particulares. Bajo esas influencias selectivas, las moléculas y sus interrelaciones se volvieron cada vez más complejas y finalmente desarrollaron las cualidades que definen las formas de vida más simples: metabolismo, crecimiento y reproducción.
Las condiciones ambientales apropiadas para la génesis de la vida probablemente incluyeron la presencia de muchos compuestos orgánicos simples en aguas primordiales. Se cree que los compuestos orgánicos simples se sintetizaron de forma natural mediante reacciones inorgánicas (es decir, no vivas) entre el amoníaco, metano, sulfuro de hidrógeno y otros compuestos que fueron abundantes en la atmósfera anterior a la vida. Estas reacciones se vieron favorecidas porque la atmósfera en ese momento era un ambiente de alta energía asociado con la radiación ultravioleta y los rayos. Los compuestos orgánicos resultantes fueron depositados en el océano primordial por lluvias, donde se concentraron progresivamente, especialmente en pozas someras en costas oceánicas, donde la tasa de evaporación habría sido alta.
Los científicos modernos han realizado experimentos de laboratorio simples que se cree que simulan esas condiciones primordiales. En matraces herméticos, las mezclas de agua y CH 4 gaseoso, NH 3 y H 2 S son provocadas por arcos eléctricos. Estos experimentos producen diversos tipos de hidrocarburos, aminoácidos (precursores de proteínas), bases nitrogenadas (precursores de ácidos nucleicos) y otros químicos orgánicos. Los científicos piensan que algo similar sucedió antes del origen de la vida en la Tierra.
Sin embargo, es un paso enorme desde la ocurrencia de condiciones ambientales apropiadas hasta la génesis espontánea de microorganismos vivos. Los científicos aún no entienden cómo ocurrió este acontecimiento trascendental —el origen de los primeros organismos—. De hecho, el límite entre los sistemas químicos complejos y los organismos vivos es algo arbitrario (por ejemplo, los virus existen en este límite). Sin embargo, existe un amplio consenso entre los científicos de que los microorganismos sí aparecieron en los océanos hace unos 3.5 mil millones de años (Cuadro 6.1). Esos primeros microorganismos fueron consumidores heterótrofos de la rica sopa de compuestos orgánicos que se habían acumulado en océanos prebiológicos a lo largo de cientos de millones de años. Los primeros microorganismos quimioautótrofos evolucionaron varios cientos de millones de años después, y los primeros fotosintéticos hace unos 2.5 mil millones de años.
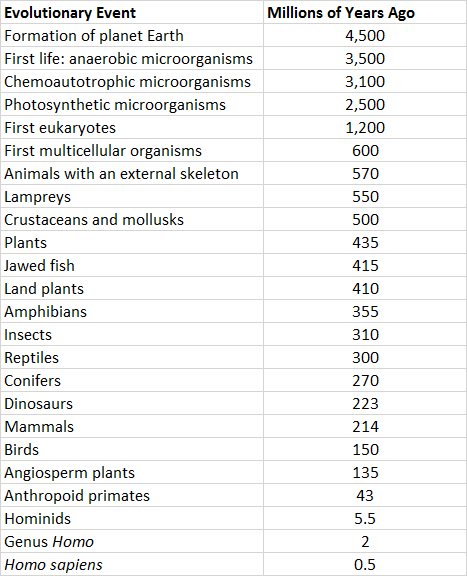
Las formas de vida más tempranas fueron los procariotas, que son organismos unicelulares que carecen de un núcleo organizado que contenga el material genético, que probablemente era ADN o ARN (ver En Detalle 6.1). Finalmente, los eucariotas (que tienen un núcleo limitado por una membrana) evolucionaron a partir de predecesores procariotas más simples.
Se cree que microorganismos más complejos, que contienen orgánulos subcelulares como mitocondrias, plastidios y cilios, han evolucionado como resultado de asociaciones simbióticas que ocurren entre diferentes especies. Según esta teoría, los microorganismos más pequeños se encapsularon dentro de otros más grandes en una simbiosis mutuamente beneficiosa (un mutualismo; ver Capítulo 9). Por ejemplo, ciertos microorganismos más pequeños pueden haber evolucionado hasta convertirse en orgánulos especializados de procesamiento de energía conocidos como mitocondrias. Otros microbios encapsulados se especializaron para capturar la luz y utilizar esa energía en la fotosíntesis, se convirtieron en cloroplastos. Las mitocondrias y cloroplastos contienen pequeñas cantidades de ADN de carácter distintivo y se cree que es residual de la antigüedad cuando estos orgánulos eran microorganismos independientes.
Los organismos multicelulares fueron la siguiente categoría principal de forma de vida en aparecer, en tiempos precámbricos tardíos (ver Tablas 3.1 y 6.1). La evolución y radiación de estos organismos complejos fue impulsada por adaptaciones fisiológicas y ecológicas asociadas con interacciones de células especializadas y, eventualmente, órganos. Los primeros organismos multicelulares eran pequeños y simples, pero estos finalmente evolucionaron hacia los organismos más grandes y complejos que ahora son prominentes en la Tierra, incluidos los vertebrados, el filo de los animales a los que pertenecen los humanos.
“Progresión” de la Vida
Todas las especies, desde las más pequeñas y simples, como las bacterias menores de 1 µm, hasta enormes ballenas azules que superan los 30 m de longitud, representan ejemplos bien adaptados y maravillosos de la diversidad de organismos. Además, en cierto sentido, los biólogos modernos creen que todas las especies vivas son igualmente “avanzadas”. Las dos razones para pensar esto son: (1) todas las especies vivas han tenido la misma cantidad de tiempo para evolucionar desde que aparecieron los primeros organismos, y (2) todas ellas están exquisitamente adaptadas para hacer frente a las oportunidades y limitaciones que presentan los entornos en los que viven.
Por supuesto, las especies también varían enormemente en su complejidad. Debemos, sin embargo, tener cuidado cuando utilizamos los términos “simple” y “complejo” en un contexto evolutivo, porque estos conceptos son difíciles de definir con precisión. De hecho, todos los organismos muestran una mezcla de rasgos, algunos de los cuales evolucionaron en la antigüedad, mientras que otros son adaptaciones más recientes. Por ejemplo, casi todos los organismos (excepto algunos virus) tienen ADN como material genético, por lo que este es un rasgo antiguo. En contraste, el vuelo en murciélagos y la inteligencia en humanos representan adaptaciones específicas que ocurrieron relativamente tarde en solo unos pocos linajes evolutivos.
El registro fósil demuestra claramente que, con el tiempo, ha habido una progresión de formas de vida en la Tierra. Los primeros organismos procariotas fueron diminutos y simples, pero a través de la evolución estos condujeron al desarrollo de microorganismos eucariotas más complejos, y así sucesivamente hasta que evolucionaron animales y plantas grandes y sumamente complejos. Este patrón evolutivo implica una clara secuencia temporal. Sin embargo, es importante entender que especies relativamente complejas y más recientes (incluidos los humanos) no representan el acme de la evolución, ni han heredado la Tierra y sus oportunidades. Más bien, todas las especies vivientes comparten este planeta abundante y su biosfera, el único lugar del universo conocido por sostener la vida.

En Detalle 6.1. Un Primer sobre Genética
Cada organismo tiene un complemento individual de información genética contenida en la disposición específica de los nucleótidos en su ADN o ARN. A continuación se presenta un breve resumen del almacenamiento y traducción de información genética.
El ADN (ácido desoxirribonucleico) transporta la información genética en casi todas las especies. En algunos virus, sin embargo, la información genética está contenida en el ARN (ver más abajo). El ADN, un ácido nucleico, consiste en secuencias lineales de solo cuatro nucleótidos: adenina, citosina, guanina y timina. Las secuencias están dispuestas como dos cadenas, que se enrollan como una doble hélice (espiral) y se mantienen unidas por enlaces de hidrógeno entre nucleótidos complementarios: adenina con timina y citosina con guanina. La información genética está incrustada en la secuencia precisa de los nucleótidos.
El ARN (ácido ribonucleico) está compuesto por una sola cadena de nucleótidos. En el ARN, el uracilo sustituye la timina del ADN. Las secuencias de nucleótidos del ARN guían la traducción de la información genética del ADN a la estructura de las proteínas (ver más adelante).
Los cromosomas están compuestos por ADN y proteínas y contienen la información genética de la célula. Los cromosomas se autoduplican, crean copias exactas de sí mismos a través del proceso de replicación (ver más abajo). Se pasa una copia exacta a cada celda hija cuando una celda se divide. Los cromosomas en las células corporales (somáticas) de plantas y animales ocurren como pares complementarios (pares homólogos). El número de pares de cromosomas varía mucho entre especies, de uno a cientos.
Los genes son regiones específicas de un cromosoma que determinan el desarrollo de un rasgo particular codificando una proteína específica durante la transcripción (ver más abajo). Debido a que los cromosomas ocurren en pares, los genes también están emparejados. Los genes ocurren comúnmente en más de una forma, cada una de las cuales se llama alelo. A menudo, un alelo es dominante (D) y el otro recesivo (r). El dominante se expresa cuando ambos alelos en un par de genes son de este tipo (DD), y también cuando ocurren alelos tanto dominantes como recesivos (rD o Dr). Los alelos recesivos se expresan sólo si ambos son de este tipo (rr). La condición en la que ambos alelos son iguales (DD o rr) se denomina homocigótica, mientras que la condición mixta (rD o Dr) es heterocigótica.
La replicación es el proceso bioquímico durante el cual se copia la secuencia de nucleótidos de cada cadena de ADN. La replicación es necesaria para que se produzca la división celular, ya que cada nueva célula requiere una copia idéntica del ADN. Durante la replicación, la doble hélice del ADN se “desbloquea”, lo que permite que los nucleótidos libres se unan de hidrógeno con los de cada cadena, produciendo moléculas de ADN nuevas pero idénticas. Si se produce un error durante la replicación, el resultado es un cambio en la información genética, que se denomina mutación.
La transcripción implica la descompresión del ADN y se realiza una cadena complementaria de ARN en una de las cadenas de ADN, de manera similar a la replicación. Entonces el ARN flota libre y el ADN se cierra de nuevo. Se pueden hacer tres tipos de ARN: (a) ARN ribosómico (ARNr), que forma pequeños cuerpos en el citoplasma llamados ribosomas; (b) ARN mensajero (ARNm), que transporta información del ADN al ribosoma; y (c) ARN de transferencia (ARNt), que se describe a continuación.
La traducción ocurre cuando el ARNm, que contiene información de una porción de una cadena de ADN, se une a un ribosoma en el citoplasma (fuera del núcleo). Allí, las moléculas de ARNt se unen a aminoácidos específicos y los transportan al ARNm en la secuencia correcta para la síntesis de una proteína en particular. (Los aminoácidos son los bloques de construcción de las proteínas. Sólo 20 aminoácidos son comunes, pero conforman la extraordinaria diversidad de proteínas que se encuentran en los organismos. Las proteínas son extremadamente importantes, principalmente como sustancias químicas estructurales y enzimas reguladoras del metabolismo). La información sobre el ARNm, copiada del ADN, determina la secuencia exacta de aminoácidos en una proteína, y por lo tanto determina su función.
La meiosis es importante en la reproducción sexual, en la que dos células “sexuales”, una de cada padre, se combinan para iniciar una nueva vida. Si esas células fueran células somáticas, cada una tendría el mismo número de cromosomas que el progenitor (el número diploide), y la progenie tendría entonces el doble del número del progenitor. Sin embargo, esto no ocurre porque las células sexuales no son diploides. En cambio, a través de la meiosis, el número de cromosomas en las células sexuales se reduce a la mitad (a haploide), por lo que la progenie tiene el mismo número de cromosomas que el progenitor.
Durante la meiosis, los cromosomas emparejados se separan, con uno de cada par yendo aleatoriamente a cada célula “sexual” hija. Justo antes de que se separen, pueden ocurrir intercambios de material genético entre los cromosomas emparejados, un fenómeno conocido como cruce. Ambos procesos incrementan la variabilidad de la información genética en las células sexuales. Cuando las células sexuales haploides (una de cada progenitor) se combinan, el resultado es una progenie diploide. Al tener cromosomas de cada progenitor, la progenie es genéticamente diferente a ellos, pero también similar. Es así como los padres pasan su información genética a sus crías.
Genotipo se refiere a la información genética única de organismos individuales, tal como se encarna en las secuencias de nucleótidos de su ADN. El genotipo único de un individuo es fijo (excepto mutaciones raras). Sin embargo, los genotipos colectivos de poblaciones y especies son bastante variables, aunque esto está restringido por el rango de variación genética entre los individuos constituyentes.
El fenotipo se refiere a la expresión real del genotipo de un individuo en términos de su desarrollo anatómico, comportamiento y bioquímica. Por ejemplo, los alelos recesivos, a menos que se encuentren en una condición homocigótica, no se expresan, aunque aparezcan en el genotipo.
Más importante aún, la expresión del potencial genético también se ve afectada por las condiciones ambientales y otras circunstancias. Por ejemplo, una planta de geranio, con un complemento fijo de información genética, puede ser relativamente alta y robusta si se cultiva en condiciones fértiles, poco pobladas y bien regadas. No obstante, si ese mismo individuo fuera cultivado en condiciones más secas, menos fértiles, más competitivas, su productividad y apariencia serían bastante diferentes. Tales patrones de crecimiento variables del mismo genotipo representan una respuesta fenotípicamente “plástica” a las condiciones ambientales. En contraste, el color de la flor de los geranios individuales (que pueden ser blancos, rojos o rosados) se fija genéticamente y no se ve afectado por sus condiciones ambientales.
La capacidad de un individuo para exhibir respuestas fenotípicamente plásticas a las variaciones ambientales está determinada genéticamente hasta cierto punto. Por lo tanto, la plasticidad fenotípica refleja tanto la capacidad genética como la expresión variable de esa capacidad, dependiendo de las circunstancias encontradas durante la vida de un individuo.
Evolución
La evolución puede definirse simplemente como cambios de base genética en poblaciones de organismos, que ocurren a lo largo de generaciones sucesivas. La evolución es una teoría de importancia crítica porque da cuenta del desarrollo de especies existentes a partir de progenitores que pueden haber sido diferentes a sus descendientes en forma y función. La realidad de la evolución es ampliamente aceptada por los científicos, tanto como la teoría de la gravedad, que explica cómo la Tierra gira alrededor del Sol así como muchos otros aspectos de la organización del universo.
Se cree que la selección natural es una causa especialmente importante del cambio evolutivo. En esencia, la selección natural predice que los organismos individuales que están mejor adaptados para hacer frente a las oportunidades o limitaciones de su entorno tendrán una mayor probabilidad de dejar descendientes. Si las ventajas adaptativas se determinan genéticamente, se pasarán a parte de la progenie, luego a las generaciones posteriores, y así sucesivamente. Este proceso dará como resultado un cambio evolutivo.
La evolución también puede ocurrir en respuesta a influencias catastróficas en poblaciones de organismos, como un incendio forestal o una inundación. Esto puede resultar en cambios más fortuitos (aleatorios) en la estructura genética de una población. Las poblaciones pequeñas están particularmente sujetas a tales influencias evolutivas no selectivas. La evolución también puede ocurrir en respuesta a elecciones hechas por humanos de rasgos deseables en ciertas especies, esto se conoce como selección cultural (o selección artificial).
Es importante entender que los organismos individuales no evolucionan. La evolución es un proceso de cambio genético de generación en generación, que ocurre en poblaciones o agrupaciones de organismos de orden superior (como especies). Esto no quiere decir que los organismos individuales no puedan mostrar respuestas variables a las condiciones ambientales. Estas respuestas están, sin embargo, limitadas por el grado de flexibilidad bioquímica, de desarrollo y conductual que permite el complemento genético de cada individuo (su genotipo). La expresión variable de la información genética de un individuo se denomina plasticidad fenotípica, pero esta respuesta a las variaciones en las condiciones ambientales no es un cambio evolutivo. Para que ocurra la evolución debe haber un cambio en la información genética colectiva de una población o especie.
La evolución puede ocurrir a diversas escalas. Los biólogos evolutivos utilizan el término microevolución para referirse a cambios relativamente sutiles que ocurren dentro de una población o especie, a menudo en solo unas pocas generaciones. Esto puede conducir a la evolución de una variedad, raza o subespecie. En contraste, la macroevolución describe la evolución de nuevas especies o grupos taxonómicos superiores, como un género, familia o clase. Los biólogos evolutivos continúan debatiendo y discutiendo los vínculos de estas escalas de evolución. ¿Los patrones de cambio macroevolutivo son simplemente los efectos acumulativos de muchos cambios microevolutivos durante largos períodos de tiempo? ¿O es la macroevolución en realidad el resultado de grandes cambios que ocurren en poco tiempo, cada uno representando un gran paso (o saltación) de la evolución? ¿O la macroevolución ocurre en ambos sentidos?
A pesar de los debates sobre muchos de sus detalles, la teoría de la evolución es un tema unificador en biología. Esto se debe a que la evolución puede ser utilizada para entender tantos fenómenos en la naturaleza. La evolución es utilizada por los científicos para explicar tanto el origen de la vida, como los extraordinarios cambios que se han producido en los organismos a lo largo de los miles de millones de años de historia biótica en la Tierra.
Relacionamiento y Descenso
Una definición biológica de especie es “un grupo de organismos que se aísla reproductivamente de otros grupos similares”. Dentro de una especie, los organismos individuales tienden a parecerse entre sí, pero lo que es más importante, pueden reproducirse entre sí y producir descendencia fértil. La incapacidad de cruzarse con éxito implica aislamiento reproductivo.
Esa especie ha evolucionado a partir de progenitores anteriores es una teoría bien establecida, ricamente respaldada por la evidencia. Algunas de las líneas argumentales más convincentes, que muestran patrones evolutivos de parentesco y descenso, se explican en las siguientes secciones.
Patrones en el Registro Fósil
Un ejemplo bien conocido de evolución que se sustenta en evidencia en el registro fósil es el del linaje equino. Uno de los primeros progenitores parecidos a caballos fue Eohippus, una criatura del tamaño de un perro que vivió hace unos 50 millones de años. Su pie tenía dos dedos fusionados y tres separados. La comparación de la morfología (estructura) de los huesos fósiles sugiere que Eohippus fue un antepasado de Mesohippus, un animal más grande que vivió hace 35 millones de años. Su pie tenía tres dedos centrales fusionados y dos exteriores libres. Una mayor evolución llevó a Merychippus, un animal algo más grande que vivía hace 20 millones de años que también tenía tres dedos fusionados y dos libres. Luego vino Pliohippus, un animal del tamaño de un poni que vivía hace 10 millones de años, que tenía los cinco dedos fusionados en una pezuña. Los caballos modernos evolucionaron hace varios millones de años y tienen un casco como el de Pliohippus. Incluyen el caballo (Equus caballus), el burro (E. asinus), el caballo salvaje mongol (E. przewalskii) y la cebra (E. burchelli).
Inferencias de las Especies Modernas
Las especies modernas muestran muchas similitudes y disimilitudes obvias que pueden usarse para agruparlas sobre la base de la relación inferida. Los primeros estudios de este tipo involucraron principalmente anatomía comparada. La investigación en animales se basó principalmente en las características de huesos, conchas, pieles y otras estructuras perdurables, mientras que los estudios de plantas involucraron en gran medida la anatomía de flores y frutos. Estudios más recientes recopilan una gama mucho más amplia de información comparativa para examinar la relación entre grupos de especies, incluyendo información sobre su comportamiento, ecología, proteínas y, más recientemente, secuencias de bases específicas del ADN. Por ejemplo, estudios que involucran ADN y proteínas sanguíneas han demostrado claramente que los humanos están estrechamente relacionados con otros grandes simios, como el chimpancé, orangután y gorila. De estos, los humanos están más estrechamente relacionados con los chimpancés —de hecho, las dos especies comparten alrededor del 99% de la información codificada en su ADN. Estas observaciones no sugieren que los humanos evolucionaron a partir de los simios modernos. Más bien, la interpretación apropiada es que los humanos y los simios vivos comparten ancestros comunes parecidos a simios.
Evolución observada
Como se acaba de describir, los patrones de parentesco y descenso pueden inferirse a partir de estudios comparativos del registro fósil y de los atributos de las especies modernas. Sin embargo, es importante entender que la evolución de una nueva especie nunca se ha observado directamente. Esto se debe a que toma mucho tiempo para que las poblaciones de organismos relacionados diverjan lo suficiente como para convertirse en nuevas especies —quizás miles a cientos de miles de años—. A pesar de esto, los biólogos no tienen ninguna duda de que las nuevas especies han estado evolucionando durante miles de millones de años, de hecho, a lo largo de la historia de la vida.
Aunque no se ha observado especiación en la naturaleza, se conocen claros ejemplos de microevolución. Estos casos proporcionan evidencia clave en apoyo de la teoría de la evolución.
Melanismo industrial
Un ejemplo de microevolución es el de la polilla salpimentada (Biston betularia) de Europa occidental. La coloración normal de esta polilla es un bronceado moteado, blanquecino. Durante el día, la polilla a menudo descansa sobre árboles cubiertos de líquenes, donde es difícil de ver contra la corteza. Este camuflaje es importante para la supervivencia de la polilla porque sus depredadores más importantes, como las aves, cazan usando la visión.
Hace aproximadamente un siglo en Inglaterra, se observó que algunas poblaciones de polillas salpicadas habían desarrollado una coloración negra, conocida como melanismo. Esto aparentemente había ocurrido en respuesta a los cambios en la corteza de los árboles locales, que habían perdido su cobertura de líquenes debido a la contaminación del aire y se habían ennegrecido por la deposición de hollín. En estas condiciones de hábitat, las polillas normales de color claro fueron altamente visibles para los depredadores y se encontraban en desventaja selectiva en comparación con las polillas más oscuras. Los estudios mostraron que el melanismo tiene una base genética, y que las polillas melánicas ocurrieron, pero eran raras, en hábitats no contaminados. Sin embargo, los individuos melánicos se volvieron dominantes en poblaciones que viven en hábitats contaminados, lo que representa un cambio genético a nivel poblacional. Este famoso ejemplo de microevolución, que también se ha demostrado en otras especies de polillas, es conocido como “melanismo industrial”.
Curiosamente, la calidad del aire ha mejorado mucho en la mayor parte de Europa occidental en las últimas décadas, en gran parte debido a la legislación de aire limpio que ha reducido el uso del carbón como fuente de energía. Como resultado, los líquenes vuelven a crecer en los árboles y las superficies de corteza tienen menos hollín. Estas recuperaciones han ido acompañadas de la reaparición de polillas de color claro en lugares donde sus poblaciones habían estado dominadas por otras melánticas, otra respuesta evolutiva a las cambiantes condiciones ambientales.
Ecotipos tolerantes a metales
En otro ejemplo de evolución observada, se encontraron varias especies de plantas creciendo en sitios en Inglaterra y Gales que estaban contaminados por desechos mineros ricos en metales. Aunque el suelo era tóxico para la mayoría de las plantas, las poblaciones de algunas especies prosperaron. Las especies más comunes fueron las gramíneas, como la hierba doblada (Agrostis tenuis). Las investigaciones mostraron que estas poblaciones locales tenían una tolerancia fisiológica de base genética a los metales tóxicos, y que diferían en este sentido de otras poblaciones de la misma especie que crecen en sitios no contaminados. Se encontró que las poblaciones adaptadas localmente, denominadas “ecotipos tolerantes a metales”, evolucionaron en tan solo varios años después de su primera exposición al suelo tóxico. (Este ejemplo fue el primero en documentarse y es famoso por esa razón. Los ejemplos canadienses de ecotipos tolerantes a metales, descubiertos posteriormente, se describen en el Capítulo 18.)

Religión y Evolución
El libro del Génesis es el primer libro de La Biblia, un texto antiguo que proporciona la base escrita para muchas de las creencias de las religiones abrahámicas (judaísmo, cristianismo e islam). La descripción de la creación divina en Génesis es la explicación escrita más antigua del origen de la vida en la Tierra, la existencia de especies y los roles y responsabilidades de los humanos en sus interacciones con el mundo natural. Sin embargo, hay algunos desacuerdos profundos entre las interpretaciones fundamentalistas del Génesis y aspectos de la teoría de la evolución. Como resultado, algunos intereses religiosos han atacado desde hace mucho tiempo la teoría de la evolución, circunstancia que se intensificó mucho después de la publicación de las ideas de Charles Darwin sobre el papel de la selección natural en la evolución (ver la siguiente sección).
Sin embargo, la ciencia y la religión no son irreconciliables. En efecto, para muchas personas, las preocupaciones físicas pertenecen al dominio de la ciencia, mientras que las espirituales pertenecen al dominio de la religión.
En todo caso, algunos grupos religiosos continúan insistiendo en una interpretación literal de la Biblia como la máxima autoridad para todo conocimiento. En particular, los creacionistas rechazan la teoría de la evolución en favor de una interpretación literal del Génesis. Afirman que el relato dado en Génesis significa que Dios creó el universo y todos los organismos vivos durante un periodo de seis días, culminando con la creación de los humanos. Los humanos fueron creados a imagen física de Dios y se les dio autoridad y poder para usar libremente los recursos de la Tierra: “Y dijo Dios, hagamos al Hombre a nuestra imagen, según nuestra semejanza; y que tengan dominio sobre los peces del mar, y sobre las aves del aire, y sobre el ganado, y sobre toda la Tierra y sobre cada cosa rastrera que se arrastra sobre la Tierra”.
Además, se instruyó a los humanos para que aumentaran sus poblaciones y explotaran la naturaleza: “Ser fructíferos, multiplicarse, y reponer la Tierra, y someterla”.
Obsérvese, sin embargo, que existe cierta controversia sobre el significado de la palabra “reponer” en este pasaje bíblico. Algunas personas lo han interpretado en el sentido de la conservación, como en para rellenar o restaurar los recursos a medida que se utilizan. Otros, sin embargo, sostienen que el significado original era “llenar”, y, en ese sentido, se refería a llenar la Tierra con humanos y sus actividades económicas. Esta última interpretación es la que suele ser aceptada en los escritos ambientales modernos que examinan la relación entre los humanos, su economía y el mundo natural.
A partir de su interpretación literal del Génesis y otros pasajes de la Biblia, los creacionistas han sacado las siguientes conclusiones relevantes para la evolución:
- La Tierra y sus especies no son antiguas porque la creación ocurrió hace apenas unos pocos miles de años.
- Las especies son esencialmente inmutables, habiendo sido creadas como entidades que no han cambiado desde su creación.
- Debido a que las especies fueron creadas individualmente, las especies existentes no descendieron de las anteriores a través de la evolución.
- Los humanos son particularmente especiales, ya que han sido creados a la imagen del Creador, no están relacionados ni descienden de ninguna otra especie.
Pero estas ideas no concuerdan con los hallazgos científicos, como se describieron en las páginas anteriores. En particular:
- El registro geológico demuestra claramente que la Tierra y el sistema solar son extremadamente antiguos, habiendo comenzado a desarrollarse hace al menos 4.5 mil millones de años. La vida también es antigua, habiéndose originado hace unos 3.5 mil millones de años. La tierra y los organismos se remontan mucho más allá de unos pocos miles de años.
- El registro fósil proporciona muchos ejemplos de grandes cambios en las características de las especies a lo largo del tiempo, al igual que los estudios de algunas especies vivas. Claramente, las especies no son inmutables. Además, el complemento existente de especies en la Tierra representa sólo una pequeña muestra de todas las que alguna vez han vivido. El registro fósil demuestra que la mayoría de las especies que evolucionaron durante la larga historia biológica de nuestro planeta están ahora extintas. Muchas de las especies extintas, familias e incluso filas no tienen descendientes vivos; todo su linaje está extinto (ver Capítulo 7).
- El registro fósil presenta evidencia clara de linajes entre grupos de organismos, lo que indica que las especies vivas han descendido de las anteriores. En casi todos los casos, las especies progenitoras están ahora extintas. Incluso hay algunos ejemplos en el registro fósil de vínculos entre grupos principales. Quizás el más famoso de estos es Archaeopteryx, una criatura de un metro de largo que vivió hace unos 150 millones de años. Tenía dientes y otros personajes dinosaurios, pero también tenía un cuerpo emplumado y podía volar. Archaeopteryx se considera un vínculo entre dinosaurios extintos y aves vivas.
- La información fósil y genética indica que los humanos descienden de especies y géneros anteriores, ahora extintos. Los registros fósiles muestran que la especie humana (Homo sapiens) deriva de un linaje evolutivo de simios antropoides. Hay algunas otras especies sobrevivientes en ese linaje, con chimpancés, y en menor grado gorilas y orangutanes, siendo los parientes vivos más cercanos de los humanos. Todos los miembros sobrevivientes de la familia de los simios descienden de progenitores ahora extintos.
Personas conocidas como creacionistas científicos también insisten en que su interpretación del Génesis es la fuente de conocimiento más confiable sobre el origen y evolución de la vida. Los creacionistas científicos han intentado explicar algunas de las discrepancias entre sus creencias y la comprensión científica actual de la evolución. Por ejemplo, algunos de ellos reconocen que la evidencia geológica y fósil sugiere que la Tierra y la vida son fenómenos antiguos y que la mayoría de las especies se han extinguido. La mayoría de los creacionistas científicos también reconocen que las especies han cambiado con el tiempo, pero solo a través de la microevolución, no están de acuerdo en que la macroevolución haya llevado al desarrollo de nuevas especies a partir de las anteriores. Por extensión, los creacionistas científicos tampoco creen que los humanos desciendan de especies anteriores de homínidos o estén relacionados con otras criaturas parecidas a simios u otros primates. Además, la teoría del creacionismo científico no abandona la noción de que, en un momento particular del pasado, Dios creó todas las especies que alguna vez han vivido en la Tierra.
La ciencia procede mediante una cuidadosa observación y pruebas de hipótesis. Pero el creacionismo científico se basa en una creencia, no en una hipótesis comprobable, respecto a una interpretación literal de la Biblia como representante de “verdad” y “conocimiento”. La mayoría de las predicciones de los creacionistas científicos no pueden ser probadas por una metodología rigurosa, pero cuando pueden serlo, son refutadas por la evidencia. En definitiva, a pesar de su nombre, el creacionismo científico no es ciencia.
Evolución por Selección Natural
Debido a que los organismos varían en su genética y fenotipos, también difieren en sus habilidades para lidiar con éxito con el estrés y las oportunidades en su entorno. Bajo ciertas condiciones, un individuo con un fenotipo particular (que está sustancialmente determinado por su genotipo) puede ser relativamente exitoso en comparación con otros que tienen diferentes genotipos y fenotipos.
En el sentido que aquí se entiende, el “éxito” de un individuo significa reproducción exitosa —tener progenie que ellos mismos van a reproducirse con éxito. Esto también se conoce como aptitud, o la contribución genética proporcionada que hace un individuo a toda la progenie en su población. Un principio central de la teoría evolutiva es que los individuos maximizan su aptitud optimizando el grado en que sus propios atributos genéticos influirán en las generaciones futuras de su especie.
Los biólogos consideran que la evolución procede principalmente por la selección natural, la cual opera cuando existe variación genética entre individuos dentro de una población, por lo que algunos de ellos están mejor adaptados para hacer frente a las condiciones ambientales imperantes. En promedio, los organismos más aptos tienen mayor éxito en la reproducción, por lo que tienen una influencia desproporcionada en la evolución de las generaciones posteriores.
La teoría de la evolución por selección natural es quizás el concepto más grande unificador en la biología moderna, ya que da contexto a prácticamente todos los aspectos del estudio de la vida. Esta teoría fue coanunciada públicamente en 1858 por dos naturalistas ingleses: Charles Darwin (1809—1882) y Alfred Russel Wallace (1823—1913). Darwin, sin embargo, había estado trabajando en aspectos de la teoría durante aproximadamente 20 años antes de su publicación, y había recopilado evidencia detallada en apoyo de la selección natural como mecanismo de evolución. La copiosa evidencia de Darwin fue reunida en el famoso libro, Sobre el origen de las especies por medio de la selección natural, publicado en 1859. Debido a este libro, Darwin se ha vinculado más estrechamente que Wallace a la teoría de la evolución por selección natural. Darwin es también el más famoso de los dos científicos, en gran parte por sus grandes contribuciones para comprender los mecanismos de la evolución. Quizás el biólogo más influyente de todos los tiempos, Darwin emprendió una gama asombrosamente amplia de proyectos de investigación sobre una gran variedad de especies y temas biológicos.
En su Origen de las Especies, Darwin resumió la selección natural de la siguiente manera: “¿Podemos dudar... que los individuos que tengan alguna ventaja, por leve que sea, sobre otros, tendrían la mejor oportunidad de sobrevivir y de procrear a su especie? Por otro lado, podemos estar seguros de que cualquier variación en el menor grado lesivo sería destruida rígidamente. Esta preservación de variaciones favorables, llamo Selección Natural”.
En un ensayo inédito que Wallace envió a Darwin para su revisión en 1858, la selección natural se expresó de una manera bastante similar: “La vida de los animales salvajes es una lucha por la existencia... en la que los más débiles... siempre deben sucumbir... dando lugar a sucesivas variaciones que parten cada vez más de la tipo original.”
La teoría de Darwin y Wallace se basó en la siguiente línea de razonamiento:
- Se sabe que la fecundidad de todas las especies es lo suficientemente alta como para que puedan sobrepoblar fácilmente sus hábitats, sin embargo esto generalmente no sucede.
- También se sabe que los recursos que las especies necesitan para sostenerse son limitados, particularmente en hábitats relativamente estables.
- Por lo tanto, ante el potencial crecimiento poblacional y los recursos limitados, debe existir una intensa competencia entre los individuos de cada especie por el acceso a las necesidades de la vida. Sólo algunos individuos logran sobrevivir a esta lucha por la existencia y reproducirse.
- Debido a que los individuos dentro de una especie son diferentes entre sí, y gran parte de esta variación es heredable, es razonable sugerir que la supervivencia en la lucha por la existencia está influenciada en parte por diferencias de habilidades determinadas genéticamente.
- Los individuos que sean más capaces tendrán más posibilidades de sobrevivir y reproducirse, y sus atributos de base genética estarán representados desproporcionadamente en las generaciones futuras.
- Durante largos periodos de tiempo, este proceso de selección natural conducirá a cambios evolutivos dentro de las poblaciones, y eventualmente a la evolución de nuevas especies.
Cuando se presentó públicamente por primera vez en 1858, la teoría de la evolución por selección natural creó sensación entre los científicos y también dentro de la sociedad. El entusiasmo y la controversia ocurrieron en gran parte porque la teoría proporcionó el primer cuerpo convincente de evidencia en apoyo de las siguientes tres nociones: (a) ocurre la evolución, (b) procede bajo influencias naturales, y (c) ha dado como resultado la gran diversidad de especies vivas.
Esta fue una visión radicalmente diferente a la del creacionismo, que fue la explicación predominante del origen de la vida y las especies a mediados del siglo XIX. Curiosamente, los escritos de Darwin no desafiaron directamente la existencia de un Creador divino. En su mayoría discutió las causas del cambio en las especies a lo largo del tiempo, y no sugirió directamente que los antepasados iniciales no habían sido creados por Dios. Las teorías modernas sobre la génesis espontánea de la vida en la Tierra se basan en una ciencia relativamente sofisticada que Darwin desconocía. Tampoco conocía los mecanismos de la genética y la herencia de rasgos.
Las extensiones modernas de la teoría de la evolución por selección natural sugieren que nuevas especies evolucionan a partir de progenitores (un proceso conocido como especiación). Se cree que esto sucede cuando las poblaciones se aíslan por barreras intervinientes como una cordillera, glaciares extensos u otras discontinuidades inhóspitas. El aislamiento es importante en la especiación porque reduce o elimina los intercambios genéticos, y por lo tanto permite que la diferenciación proceda de manera más efectiva. Las poblaciones aisladas que experimentan diferentes condiciones ambientales están sujetas a diferentes presiones de selección y pueden evolucionar de maneras diferentes. Eventualmente, puede haber suficiente cambio evolutivo como para que las poblaciones ya no puedan cruzarse con éxito, aunque se reúnan espacialmente. En ese punto, las poblaciones han logrado el aislamiento reproductivo, y así se han vuelto especies estrechamente relacionadas pero diferentes.
También se piensa que la especiación ocurre de una manera más lineal, como cuando las especies progenitoras evolucionan gradualmente con el tiempo en respuesta a los cambios en las condiciones ambientales. Eventualmente, las especies ancestrales pueden extinguirse, pero otras nuevas evolucionadas a partir del linaje progenitor pueden sobrevivir para continuar la cadena evolutiva.
La Importancia de la Genética
El conocimiento de la genética en la época de Darwin se basó en una comprensión altamente incompleta de cómo se transmiten los rasgos de un organismo a su descendencia. Una teoría popular, la “herencia de rasgos adquiridos”, se basó en la observación de que la morfología, el comportamiento y/o la bioquímica de los organismos individuales podrían variar en respuesta al cambio ambiental. Según la teoría, estas respuestas plásticas a las condiciones ambientales podrían transmitirse a la progenie de un individuo. Por ejemplo, durante períodos de sequía o intensa competencia por la alimentación, los ancestros individuales de las jirafas de cuello corto podrían haber estirado el cuello hasta donde pudieron para alcanzar el escaso follaje más alto en los árboles, resultando en el desarrollo de un cuello más largo. El largo cuello se habría pasado a la progenie de la jirafa, quien la desarrolló aún más. Finalmente, las poblaciones desarrollaron el familiar cuello largo de las jirafas modernas.
Sin embargo, la selección natural sugiere un mecanismo diferente de este cambio evolutivo: dentro de las poblaciones de jirafas de cuello corto existía una variación genéticamente determinada en la longitud del cuello entre los individuos. Debido a que las jirafas de cuello largo eran más capaces de encontrar comida, tenían más probabilidades de sobrevivir y reproducirse. Esto significó que más de la siguiente generación tenían el rasgo de cuello largo, y esta característica anatómica se hizo cada vez más prominente en la población en evolución.
Las observaciones y experimentos modernos han demostrado que los “rasgos adquiridos” son solo una manifestación de la plasticidad fenotípica. No hay evidencia de que puedan fijarse genéticamente en un individuo y transmitirse a su descendencia. En contraste, la ciencia de la genética ha aportado pruebas convincentes en apoyo de la teoría de la evolución por selección natural. Se han descubierto los mecanismos bioquímicos que determinan el genotipo de un organismo individual y cómo algunas de sus características pasan a la progenie. El tema es bastante complicado y no se puede tratar aquí con mucha profundidad. Sin embargo, es útil examinar los experimentos clave que primero sugirieron la existencia de genes.
Esta investigación fue realizada por Gregor Mendel (1822—1884), un científico austríaco (y monje) que desarrolló importantes ideas sobre la herencia a través de experimentos de cría con el guisante de jardín (Pisum sativum). Mendel estaba interesado en producir híbridos de guisantes, lo que implica cruzar dos plantas parentales, cada una con rasgos distintivos. Investigaciones previas habían demostrado que ciertos rasgos se fijaban en variedades cultivadas de guisantes, incluyendo el color de la flor (blanca o púrpura) y si las semillas tienen un pelaje arrugado o liso. En total, Mendel trabajó con 32 rasgos de este tipo. Las flores de guisante son bisexuales, contienen partes tanto femeninas (pistilo, que contienen los óvulos) como macho (anteras, que contienen polen). Estos son compatibles dentro de un mismo individuo, por lo que puede ocurrir la autofecundación. Sin embargo, Mendel experimentó mediante la fertilización cruzada de padres seleccionados, produciendo hibridaciones conocidas.
En cada experimento, Mendel cruzó dos variedades endogámicas en las que ciertos rasgos “se reproducían verdaderos” (eran homocigóticos, como por ejemplo para un color de flor blanca o púrpura). La progenie (primera generación) eran todas iguales: todas tenían flores moradas. Sin embargo, los cruces entre las plantas de primera generación arrojaron una relación de aproximadamente tres flores moradas y una flor blanca en la segunda generación. Esto se ajusta a la predicción de cruces de dos generaciones entre dos líneas homocigóticas, de la siguiente manera:
- Representar la variedad púrpura original como AA. Este rasgo es dominante sobre el rasgo de flor blanca (llamado recesivo).
- Representar la variedad blanca original como aa.
- Cuando se cruzan las dos plantas, la progenie de primera generación tiene flores moradas pero son heterocigóticas (Aa).
- Un cruce entre las plantas de primera generación arroja cuatro resultados posibles: AA, Aa, aA y aa. Cada uno es igualmente probable. Debido a que A es dominante a, la progenie AA, Aa y aA tienen flores moradas. Sólo aa tiene flores blancas. Por lo tanto, la relación esperada de púrpura a blanco entre la progenie de segunda generación es 3:1.
La conclusión más importante que surgió del trabajo de Mendel fue que la herencia de información genética ocurre en forma “particulada” (a la que ahora nos referimos como genes), involucrando a menudo alelos dominantes y recesivos. La herencia no es una condición de mezcla, en el ejemplo que se acaba de describir, un cruce de plantas de guisantes de flores moradas y blancas no produce progenie de un color intermedio. Por lo tanto, el color de la flor y muchos otros rasgos son unidades discretas que permanecen intactas durante la herencia y se expresan o no en la progenie.
Mendel publicó por primera vez sus resultados en 1865 en una revista relativamente oscura. Como resultado, el trabajo fue desconocido para la corriente principal de la ciencia durante muchos años. Sin embargo, la obra de Mendel fue redescubierta y republicada en 1900 y rápidamente se convirtió en la base de las teorías modernas de la herencia genética.
El trabajo de Mendel y el posterior florecimiento de la ciencia de la genética han sido extremadamente importantes en biología y en el desarrollo de la teoría moderna de la evolución. Esto se debe a que la genética permite una explicación racional de la herencia como mecanismo por el cual los rasgos genéticamente fijados pueden transmitirse a la descendencia. Investigaciones posteriores han encontrado que pueden surgir nuevos genotipos a través de diversos mecanismos, como la hibridación, el poliploidismo (un aumento espontáneo en el número de cromosomas) y las mutaciones. La variación genética es, por supuesto, el menú de posibilidades entre las que puede elegir la selección natural para que pueda ocurrir la evolución adaptativa.
Es importante reconocer que mucha información genética en un individuo no parece codificar enzimas funcionales u otras proteínas, y por lo tanto no codifica para rasgos que podrían seleccionarse a favor o en contra. Por su neutralidad con respecto a la selección natural, a este tipo de material genético se le denomina a veces “ADN basura”. Sin embargo, podemos ser ignorantes de otros roles que el llamado ADN basura puede desempeñar en el funcionamiento del genoma.
Cuestiones Ambientales 6.1. Organismos
genéticamente modificados Los organismos genéticamente modificados (OGM) son un tema muy controvertido. Pero ¿qué, exactamente, se entiende por el término?
En sentido estricto, los OGM son organismos cuyo genotipo ha sido influenciado por la intervención humana. Pero la gente ha estado haciendo esto desde hace muchísimo tiempo. Ya hace unos 10 mil años, cuando las personas comenzaron a cultivar otras especies como cultivos, criaban selectivamente plantas y animales individuales que tenían rasgos favorables (ver Capítulo 10 para una explicación de la evolución sociocultural, incluido el desarrollo temprano de la agricultura). Esta “selección artificial” condujo rápidamente a la evolución de variedades de cultivos que fueron más receptivas al manejo y tuvieron mayores rendimientos que sus progenitores silvestres. En este sentido, casi todas las especies domesticadas de plantas, animales y microorganismos que se cultivan como fuentes de alimento, material o energía son “organismos genéticamente modificados”. Se produjeron utilizando métodos convencionales de mejoramiento selectivo, un proceso que no es muy polémico.
Más recientemente, sin embargo, se han utilizado nuevas técnicas en biotecnología, específicamente en biología molecular, para crear nuevas modificaciones genéticas de organismos. Estas técnicas permiten a los biólogos insertar selectivamente porciones del ADN de una especie en el genoma de otra especie. Se trata de un tipo fundamentalmente diferente de modificación genética que la cría selectiva, y debería denominarse más adecuadamente como modificación transgénica, o como bioingeniería recombinante. Existen beneficios potenciales para este tipo de modificación genética de especies de cultivos, incluyendo el desarrollo de variedades que sean resistentes a enfermedades o plagas y que requieran menos fertilizante o pesticida. A pesar de estos aparentes beneficios, existe controversia sobre la biotecnología transgénica y el uso comercial de OGM, en gran parte por los siguientes temas:
- ¿Deberían los científicos estar interfiriendo con la base misma de la vida, la genética de las especies, mediante el uso de métodos de “ingeniería” genética que normalmente no existen en la naturaleza?
- ¿Los organismos transgénicos novedosos representan “nuevas” variedades de vida diseñada y manufacturada que son apropiadas para patentar y usar con fines comerciales? (De hecho, diversas resoluciones legales han afirmado que esto se puede hacer, y algunos cultivos transgénicos se han vuelto extremadamente rentables para los propietarios de las patentes).
- ¿Los riesgos ecológicos importantes están asociados con el cultivo de organismos transgénicos? Debido a que muchas incógnitas biológicas y ecológicas están asociadas con esta práctica, pueden surgir “sorpresas” de la liberación de estos organismos al medio ambiente, incluyendo daños imprevistos a cultivos, especies silvestres y ecosistemas naturales.
Se trata de cuestiones polémicas y cautelares, y la controversia no se resuelve. En algunos casos, las liberaciones ilegales de productos OGM han sido realizadas por intereses privados, circunstancia que refleja debilidad en los mecanismos de control regulatorio (Clapp, 2008). Sin embargo, algunos productos OGM han sido ampliamente comercializados y ahora se utilizan de manera rutinaria. Por ejemplo, se han desarrollado variedades transgénicas transgénicas de soya y canola para ser resistentes al glifosato, lo que permite que este herbicida sea utilizado en esos cultivos. Esta práctica resulta en beneficios para los agricultores por la reducción de los costos de energía y maquinaria necesaria para controlar las malas hierbas. De manera similar, se han desarrollado variedades transgénicas de maíz (maíz) que contienen ADN modificado de la bacteria insecticida Bacillus thuringiensis. Esto proporciona resistencia a plagas importantes de insectos y permite a los agricultores usar menos insecticida. Estos y otros cultivos transgénicos ahora se cultivan ampliamente en América del Norte (aunque están prohibidos en otros países, incluyendo la mayor parte de Europa y Brasil), pero se sabe relativamente poco sobre los riesgos biológicos y ecológicos que pueden surgir cuando sus factores transgénicos escapan a las plantas silvestres.
Mecanismos adicionales de evolución
Si bien la selección natural es el mecanismo más importante de la evolución, no es el único. Por ejemplo, la selección artificial implica la cría deliberada de plantas, animales y microorganismos para mejorar ciertos rasgos que los humanos ven como deseables. La selección artificial tiene evidentes paralelismos con la selección natural, ya que los organismos individuales con rasgos específicos de base genética experimentan un mayor éxito en la reproducción, por lo que quedan sobrerrepresentados en las generaciones posteriores. Sin embargo, los rasgos que se favorecen en la selección artificial pueden no ser adaptativos en el mundo natural. Además, debido a que se puede controlar la cría de genotipos deseados, el cambio evolutivo ocurre mucho más rápidamente bajo selección artificial que bajo selección natural.
Por ejemplo, el maíz (o maíz, Zea mays) es un cultivo importante que, a través de la selección artificial, ahora difiere enormemente de su progenitor silvestre más cercano, un pasto mexicano conocido como teosinte (Euchlaena mexicana). La selección artificial ha provocado muchos cambios evolutivos en el maíz. Por ejemplo, la cabeza fructífera (que consiste en la mazorca y las semillas) es mucho más grande que en los ancestros silvestres del maíz; las semillas tienen diferente coloración; las semillas se implantan de forma segura sobre la mazorca para que no se dispersen antes de cosechar; el fruto maduro se envuelve firmemente dentro de las hojas que encierran conocidas como brácteas, nuevamente para prevenir pérdidas previas a la cosecha; y hay respuestas de crecimiento vigorosas a la aplicación de fertilizantes, control de malezas y otras prácticas de cultivo. Además, sin la intervención de los humanos a través del cultivo, es probable que el maíz se extinga en solo unas pocas generaciones. Esto se debe en parte a que la selección artificial ha hecho que sus semillas sean prácticamente incapaces de desprenderse de la mazorca, que en cualquier caso está fuertemente unida en brácteas frondosas. Por lo tanto, la dispersión de semillas sin ayuda es casi imposible.
Todas las especies domesticadas de plantas, animales y microbianos han sido sometidas a selección artificial por rasgos deseables. A veces, sin embargo, la selección artificial procede en direcciones extrañas, con el fomento de rasgos genéticos que se consideran deseables por razones estéticas más que prácticas. Por ejemplo, los criadores orientales de peces mascotas han producido algunas variedades asombrosas de peces de colores (Carassius auratus) y koi (una variedad de carpas de color dorado, Cyprinus carpio). Estas variedades, a menudo con formas y comportamientos grotescos, serían rápidamente eliminadas en una población silvestre pero son apreciadas como especímenes inusuales y valiosos por los aficionados a estas mascotas acuáticas. Se podrían hacer comentarios similares sobre curiosas variedades de gatos, perros, palomas y muchos tipos de plantas hortícolas.
La evolución también puede ocurrir a través de un proceso conocido como deriva genética, o cambios aleatorios en las frecuencias de los genes que ocurren en poblaciones pequeñas y aisladas. Tales poblaciones a menudo existen en las islas, o pueden ser creadas a través de una reducción catastrófica de una población mayor debido a una enfermedad, perturbación o algún otro factor. La base genética relativamente pequeña de poblaciones pequeñas a veces se llama “cuello de botella”. La evolución posterior se basa en la variación genética restringida de solo unos pocos individuos, que puede reducirse aún más a través de los efectos de la endogamia (reproducción entre individuos estrechamente relacionados, como hermanos). Dada la cantidad restringida de variación genética, la evolución de una población pequeña puede proceder de manera muy diferente a la de una población mayor.

Conclusiones
La Tierra es el único lugar en el universo que definitivamente se sabe que sustenta la vida y los ecosistemas. Se piensa que la vida surgió espontáneamente hace al menos 3.5 mil millones de años, por la existencia de condiciones ambientales apropiadas para su génesis. Desde ese origen, se han producido profundos cambios en la morfología y funcionalidad de los organismos a través de un proceso conocido como evolución. La evolución puede definirse simplemente como cambios en la composición genética de poblaciones y especies a lo largo del tiempo (los organismos individuales no evolucionan). Si bien la evolución ha influido en la vida en la Tierra desde que comenzó, existe controversia sobre los mecanismos del proceso. Casi todos los biólogos creen que la selección natural ha sido la causa más importante del cambio evolutivo, pero algunos piensan que las catástrofes geológicas (como los impactos de meteoritos del planeta o las intensas erupciones volcánicas) también han tenido una gran influencia.
Preguntas para revisión
- ¿De qué manera la evolución de los organismos, especialmente los capaces de fotosíntesis, ha dado lugar a cambios importantes en la química del ambiente?
- ¿Qué evidencia sustenta la teoría de la evolución?
- ¿Cómo es la selección natural un mecanismo de evolución? ¿Cuáles son otros medios por los cuales puede ocurrir la evolución?
- ¿Cómo resulta la selección artificial en la evolución de las especies domesticadas?
Preguntas para Discusión
- ¿Por qué muchos biólogos son reacios a describir ciertas especies como “más avanzadas” o “más evolucionadas” que otras?
- ¿Cómo podrían las condiciones ambientales experimentadas durante su vida haber afectado su propio desarrollo? Relacionar su respuesta con el fenómeno de la plasticidad fenotípica.
- ¿Por qué es importante el conocimiento de la genética para comprender los procesos evolutivos?
- ¿Crees que hay suficiente evidencia científica en apoyo de la idea de generación espontánea de vida para que reemplace las nociones basadas en la fe de la creación divina?
Explorando problemas
- Se le ha pedido que participe en un debate sobre la génesis y evolución de la vida. ¿Qué tipo de evidencia utilizaría para apoyar la teoría de que la vida comenzó a partir de la materia inanimada hace miles de millones de años? ¿Qué evidencia respalda la teoría de que los humanos evolucionaron a partir de ancestros anteriores que ahora están extintos?
Referencias citadas y lecturas adicionales
Bengtson, S. (ed.). 1995. Vida Temprana en la Tierra. Prensa de la Universidad de Columbia, Nueva York, NY.
Clapp, J. 2008. Liberaciones ilegales de OMG y responsabilidad corporativa: Cuestionando la efectividad de las medidas voluntarias. Economía Ecológica, 66:348-358.
Cowen, R. 2000. Historia de la Vida. 5ta ed. Blackwell Scientific, Londres, Reino Unido.
Coyne, J.A. y H.A. Orr. 2004. Especiación. Asociados Sinauer, Sunderland, MA.
Darwin, C.D. 1859. Sobre el origen de las especies por medio de selección natural, o la preservación de razas favorecidas en la lucha por la vida. Murray, Londres, Reino Unido.
Dawkins, R. 1996. El relojero ciego. WW. Norton, Nueva York.
Eldredge, N. 2000. El patrón de la evolución. W.H. Freeman & Co., San Francisco, CA.
Gould, S.J. (ed.). 1993. Libro de la Vida. Hutchinson, Londres, Reino Unido.
Hall, B.K. y B. Hallgrímsson. 2013. La evolución de Strickberger. 5ta ed. Jones & Bartlett Learning, Bostón, MAMÁ.
Horgan, J. 1991. En el Comienzo... Scientific American, 264:116-25.
Klug, W. S. y M.R. Cummings. 2011. Conceptos de Genética. 10a ed. Prentice Hall, Upper Saddle River, Nueva Jersey.
Lewin, R. 1991. Hilo de la vida: El Smithsoniano mira la evolución. Smithsonian Books, Washington, DC.
Mayr, E. 1982. El crecimiento del pensamiento biológico. Harvard University Press, Cambridge, MA. Consejo Nacional de Investigaciones. 1990. La búsqueda de los orígenes de la vida. Prensa de la Academia Nacional, Washington, DC.
Números, R.L. 1993. Los creacionistas: La evolución del creacionismo científico. Prensa de la Universidad de California, Berkeley, CA.
Raven, P.H., G.B. Johnson, K.A. Mason, y J. Losos. 2013. Biología. 10a ed. McGraw-Hill, Columbus, OH.
Ridley, M. 2003. Evolución. 3ª ed. Blackwell Science, Boston, MA.
Shapiro, R. 1986. Orígenes: Una guía escéptica para la creación de vida en la Tierra. Simon & Schuster, Nueva York, NY.