
10.2: Reacciones de fosforilación - enzimas quinasa
- Page ID
- 2385

10.2A: ATP - el donador principal de los grupos de fosforilogrupos de fosforilo
Enzimas llamadas quinasas catalizan la transferencia de grupos fosforilo a las moléculas orgánicas. El origen del grupo fosforilo en la mayoría de reacciones de fosforilación es una molécula llamada adenosina trifosfato, abreviada ATP.grupos fosforilo a las moléculas orgánicas. El origen del grupo fosforilo en la mayoría de reacciones de fosforilación es una molécula llamada adenosina trifosfato, abreviada ATP.
![atp[1][7].gif](https://espanol.libretexts.org/@api/deki/files/16759/atp%255B1%255D%255B7%255D.gif?revision=1&size=bestfit&width=341&height=180)
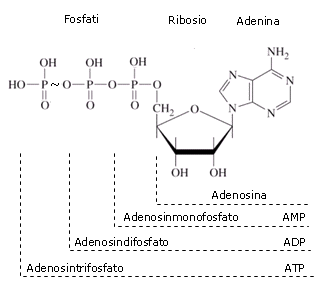
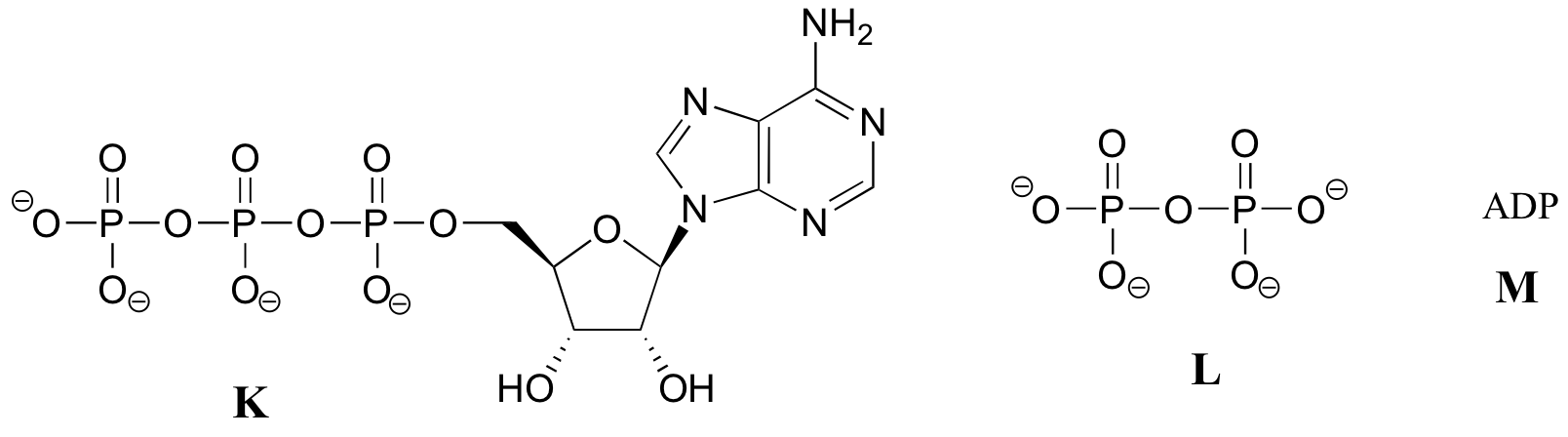
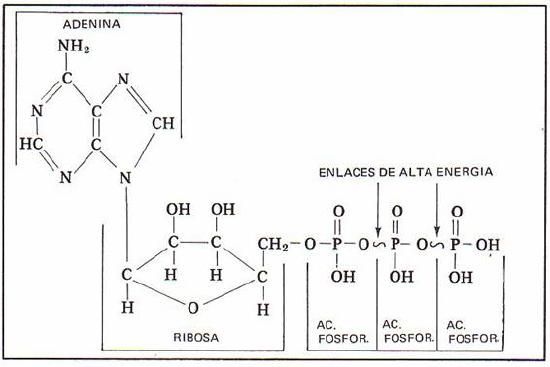
Observe que hay tres partes esenciales en esta molécula: un nucleósido de adenina, un azúcar de cinco carbonos (ribosa), y un grupo trifosfato. Los tres fosfatos se designan por letras griegas a, b, y g. Difosfato de adenosina (ADP) y monofosfato de adenosina (AMP) también son actores importantes en las reacciones de este capítulo.
Verá ATP, ADP, AMP y abreviados de muchas maneras diferentes, en este texto y en toda la literatura bioquímica. Por ejemplo, las tres estructuras de abajo son abreviaciones de ATP:
:
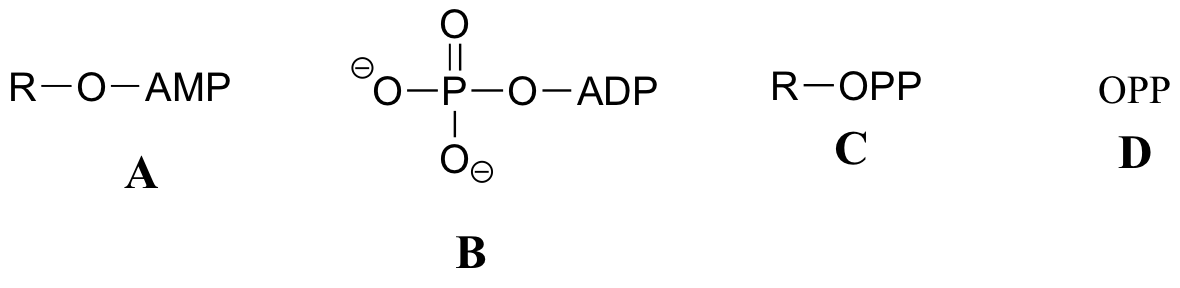
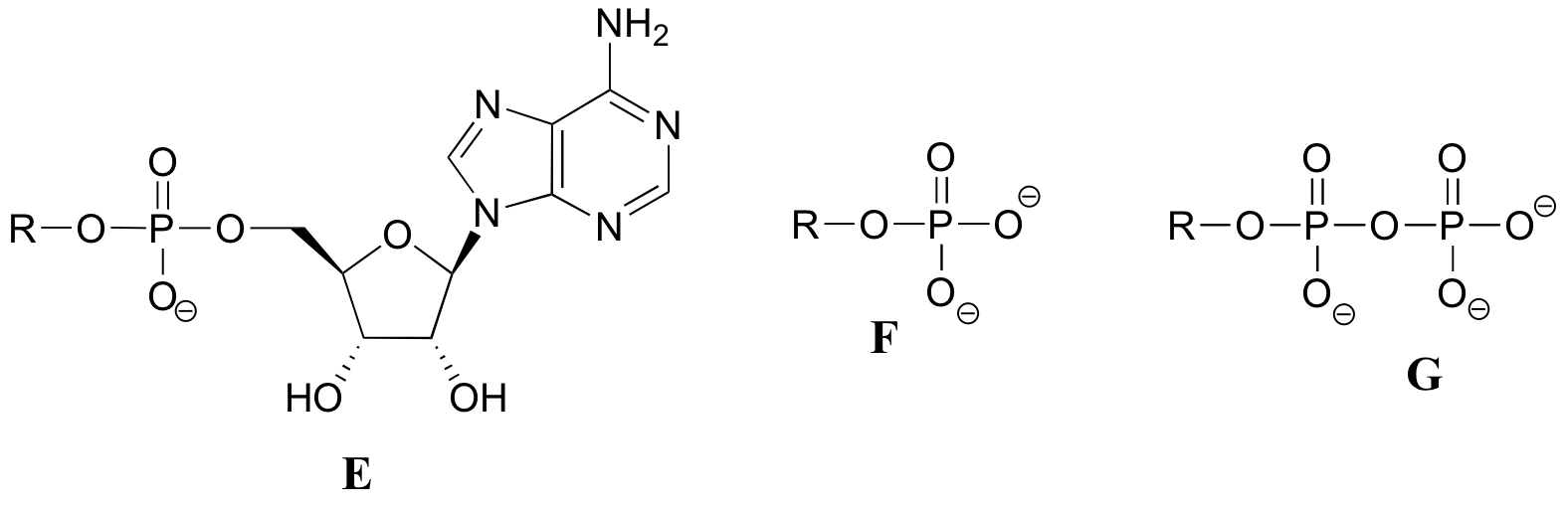
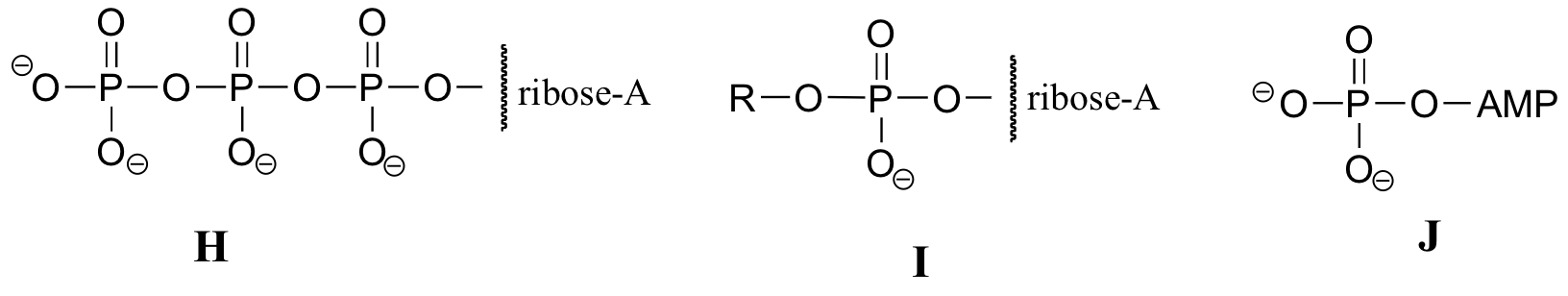
El siguiente ejercicio servirá de práctica en el reconocimiento de diferentes abreviaturas para las moléculas biológicas que contienen grupos fosfato.grupos fosfato.
Ejercicio 10.2
A continuación se presentan una serie de representaciones, denominadas A-S, de moléculas que contienen grupos de fosforilo. Se utilizan diferentes abreviaturas. Organice A-S en grupos de dibujos que representan la misma molécula, usando diferentes abreviaturas (o ninguna abreviatura en absoluto).grupos de fosforilo. Se utilizan diferentes abreviaturas. Organice A-S en grupos de dibujos que representan la misma molécula, usando diferentes abreviaturas (o ninguna abreviatura en absoluto).

Cuando hablamos de la energía ‘gastada’ en una molécula de ATP, lo que queremos decir es que un grupo fosforilo es transfiere de el ATP a alguna otra molécula aceptadora, haciendo que la molécula aceptadora más reactiva. Por ejemplo, en muchas reacciones de fosforilo es transfiere de el ATP a alguna otra molécula aceptadora, haciendo que la molécula aceptadora más reactiva. Por ejemplo, en muchas reacciones de transferencia de fosforilo (como la fosforilación de la glucosa, la cual usamos como un ejemplo en la ejemplo en la fosforilo (como la fosforilación de la glucosa, la cual usamos como un ejemplo en la ejemplo en la sección 10.1D) el fosfato gamma (γ) de ATP se transfiere a un aceptor orgánico, liberando ADP
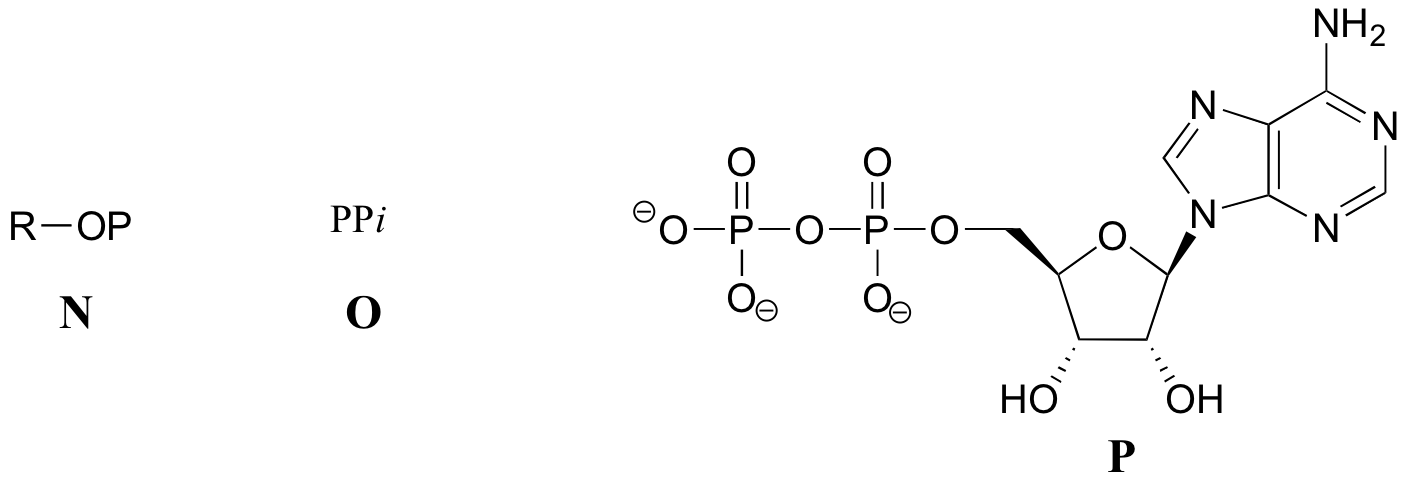
En otras reacciones, la base, ribosa y el fosfato alfa se transfiere a la molécula orgánica para formar un educto orgánico-AMP, mientras mientras pirofosfato inorgánico (PPi) se libera. mientras pirofosfato inorgánico (PPi) se libera.
De vez en cuando, los grupos fosfato beta y gamma se transfieren juntos, con la liberación de AMP.
AMP.png/=ATP_-%253E_AMP.png" fileid="16757" width="431px" height="153px" />
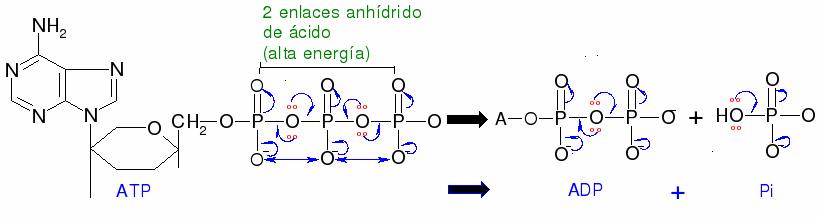
En todas estas reacciones, una molécula orgánica relativamente estable se transforma en un producto fosforilado de energía más alto. Este producto activado puede pasar a reaccionar de una manera que su contraparte más pasar a reaccionar de una manera que su contraparte más estable, no fosforilada no podía - grupos de fosforilo, como sabemos, son mejores grupos salientes en las reacciones de sustitución nucleofílica que en los grupos hidroxilo de los alcoholes. A pesar de que la conversión de un compuesto de energía más baja a una energía mas alta es, por sí mismo, un proceso termodinámicamente de grupos de fosforilo, como sabemos, son mejores grupos salientes en las reacciones de sustitución nucleofílica que en los grupos hidroxilo de los alcoholes. A pesar de que la conversión de un compuesto de energía más baja a una energía mas alta es, por sí mismo, un proceso termodinámicamente de cuesta arriba, la reacción global de transferencia de fosforilo es termodinámicamente cuesta abajo, porque la conversión de ATP en ADP o AMP - la ruptura de un vínculo anhídrido de fosfato - libera una gran cantidad de gran cantidad de energía. En otras palabras, la energía almacenada en el enlace anhídrido de fosfato de ATP se ha "gastado" para crear una molécula activada (de mas energía). Cuando AMP o ADP se convierte de nuevo en ATP, la energía de moléculas de combustible (o de la luz solar) es necesario para volver a formar el enlace anhídrido de alta energía (este proceso es objeto de discusión más adelante en esta sección)
La explicación de por qué los vínculos anhídridos de fosfato en ATP son tan energéticos reside principalmente en el concepto de separación de cargas. Recuerde de la sección 10.1 que ATP, a fisiológico pH de ~ 7, está casi completamente ionizado con una carga total de cerca de -4. Cuando uno de los dos enlaces anhídrido se rompe, las cargas negativas de los grupos fosfato son capaces de separarse y eliminar algunas de la repulsión de la misma carga que existía en ATP. Una manera de representar esto es como una espiral que salte abierta.eliminar algunas de la repulsión de la misma carga que existía en ATP. Una manera de representar esto es como una espiral que salte abierta.grupos fosfato son capaces de separarse y eliminar algunas de la repulsión de la misma carga que existía en ATP. Una manera de representar esto es como una espiral que salte abierta.eliminar algunas de la repulsión de la misma carga que existía en ATP. Una manera de representar esto es como una espiral que salte abierta.
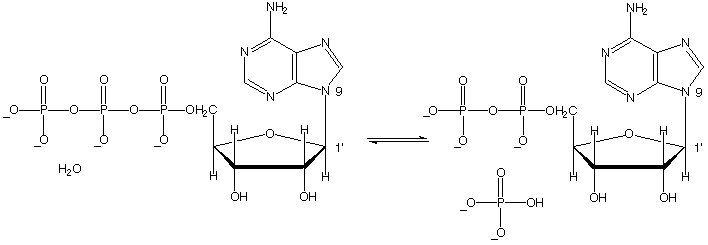
Otra razón tiene que ver con la energía de solvatación por el agua. Cuando el fosfato gamma del ATP es transferido a un alcohol, por ejemplo, las moléculas de agua alrededor son capaces de formar más enlace de hidrógeno con los productos (ADP y el fosfato orgánico) que las posibles con ATP y alcohol. Estas interacciones adicionales de solvatación estabilizar los productos de la reacción de fosforilación en relación con los compuestos iniciales.
Usted aprenderá más sobre el papel termodinámico de ATP en las vías metabólicas si toma una clase de bioquímica - lo más importante a entender a este punto es que, debido a la energía almacenada en sus enlaces anhídrido de fosfato, ATP es un poderoso grupo donante de fosforilo, y se utiliza como tal en muchas reacciones bioquímicas importantes. Algunos ejemplos son discutidos en el resto de esta sección.
10.2B: Monophosphorylation of alcohols
Recall that almost all biomolecules are charged species, which 1) keeps them water soluble, and 2) prevents them from diffusing across lipid bilayer membranes. Although many biomolecules are ionized by virtue of negatively charged carboxylate and positively charged amino groups, the most common ionic group in biologically important organic compounds is phosphate - thus the phosphorylation of alcohol groups is a critical metabolic step. In alcohol phosphorylations, ATP is almost always the phosphate donor, and the mechanism is very consistent: the alcohol oxygen acts as a nucleophile, attacking the gamma-phosphorus of ATP and expelling ADP (look again, for example, at the glucose kinase reaction that we first saw in section 10.1D).
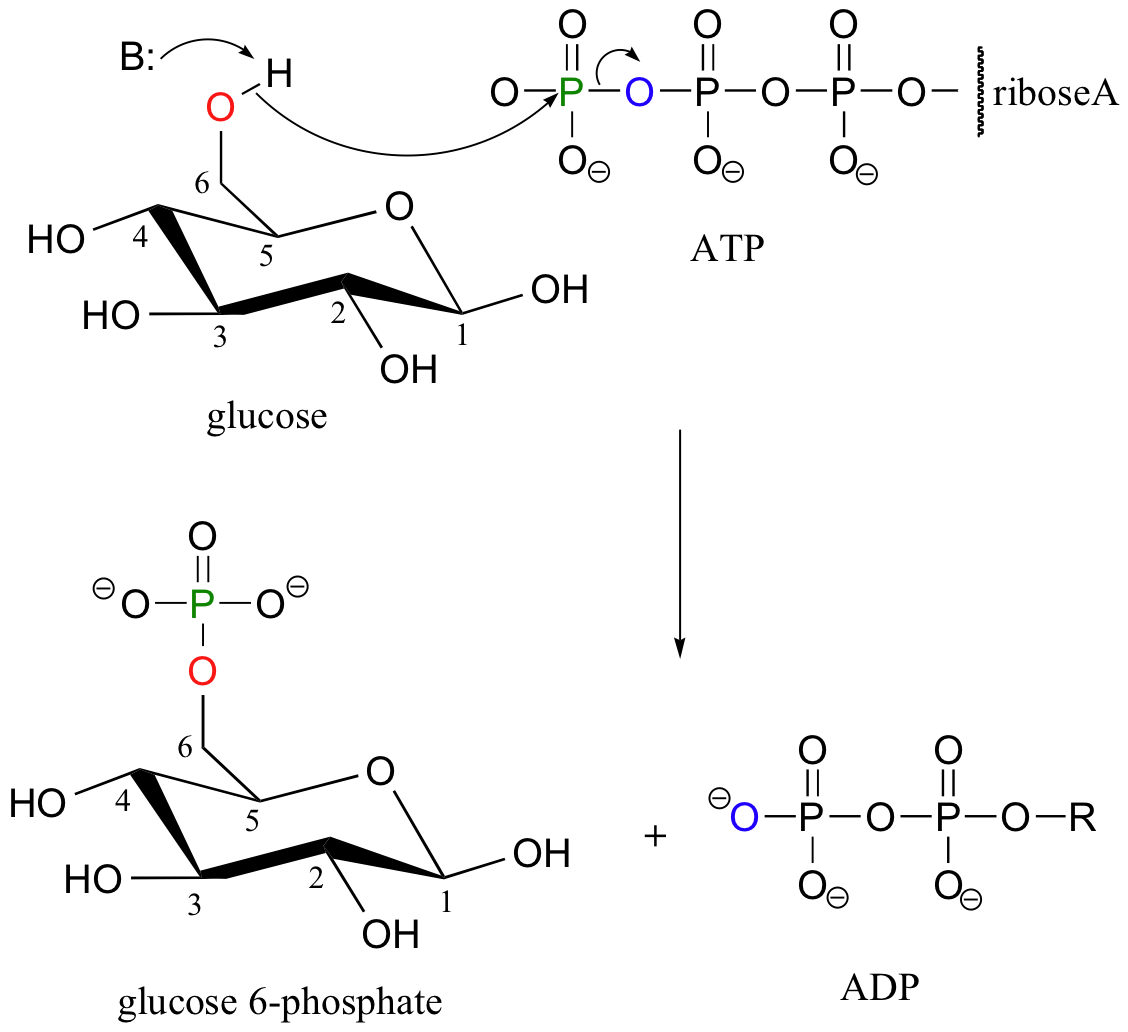 + B-H
+ B-H
The glucose kinase reaction is the first step in glycolysis, a metabolic pathway in which the 6-carbon sugar glucose is broken down into two 3-carbon fragments called pyruvate. The third step of glycolysis also a kinase reaction: this time, it is the hydroxyl group on carbon #1 of fructose-6-phosphate that is phosphorylated (step 2 of glycolysis is the isomerization of glucose-6-phosphate to fructose-6-phosphate, a reaction we will study in section 13.2A) Once again, ATP is the phosphate donor in the fructose-6-phosphate kinase reaction:
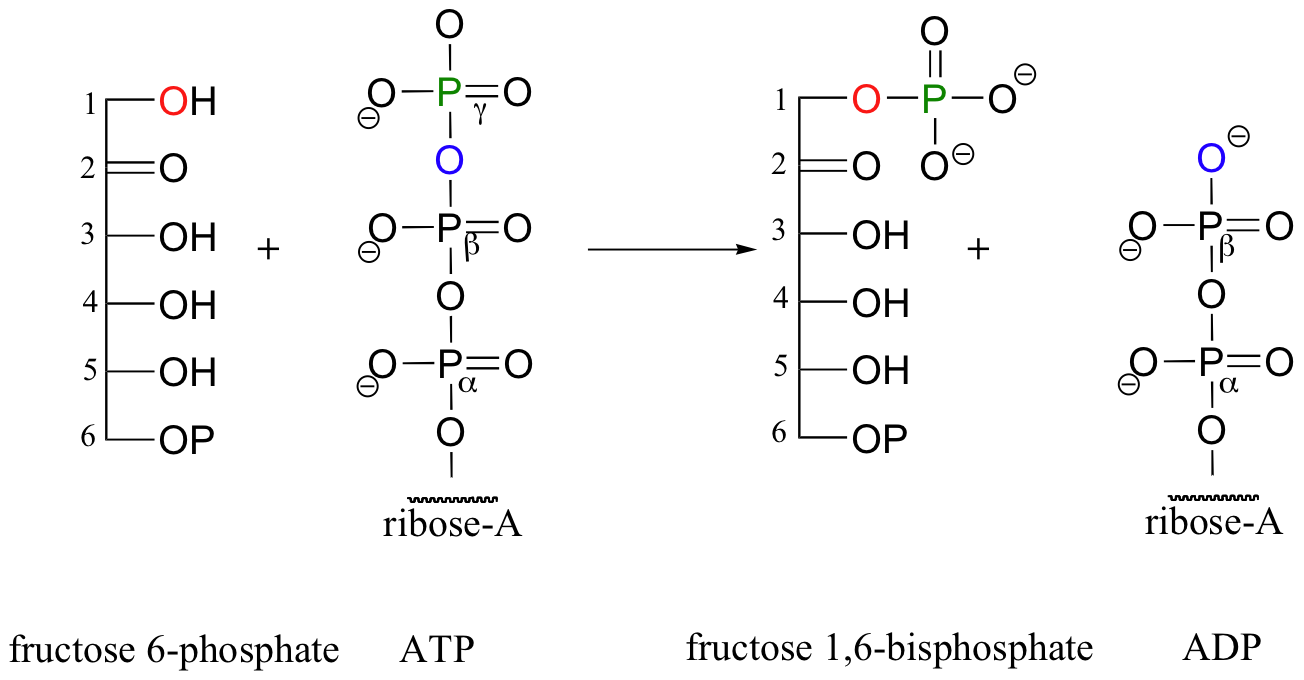
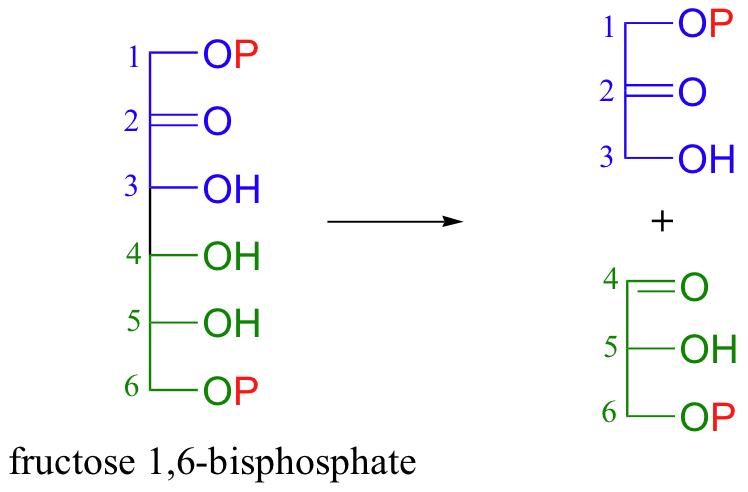
Now, when the 6-carbon sugar breaks into two 3-carbon pieces, each piece has its own phosphate group (the carbon-carbon bond-breaking step is a reaction that we will learn about in section 13.3C).
The biological activity of many proteins is regulated by means of a very similar phosphorylation reaction catalyzed by protein kinases. In these reactions, the side chain hydroxyl groups on serine, threonine, and tyrosine residues of certain proteins are modified with the gamma phosphate from ATP.

Notice the new " ATP in, ADP out" notation used in this figure, showing that ATP is converted to ADP in the course of the reaction. From here on, we will frequently use this shorthand convention to indicate when common molecules such as ATP, water, or phosphate are participants in a reaction, either as reactants or products.
Example 10.3
Example 10.4
10.2C: Diphosphorylation of alcohols
We have just seen how alcohol groups can be converted to monophosphates using ATP as the phosphate donor. In some biochemical pathways, the next step is the addition of a second phosphate group to form a diphosphate. In the early stages of the biosynthesis of ‘isoprenoid’ compounds such as cholesterol, for example, two phosphates are added sequentially to a primary alcohol group on an intermediate compound called mevalonate. The first phosphorylation is essentially the same as the reactions described in part B of this section.
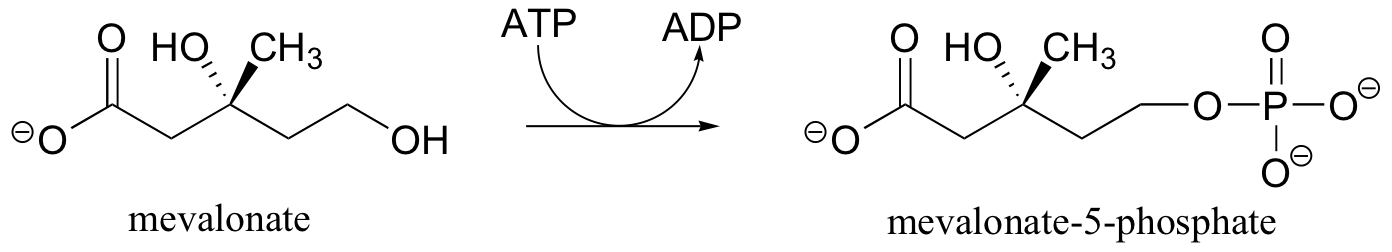
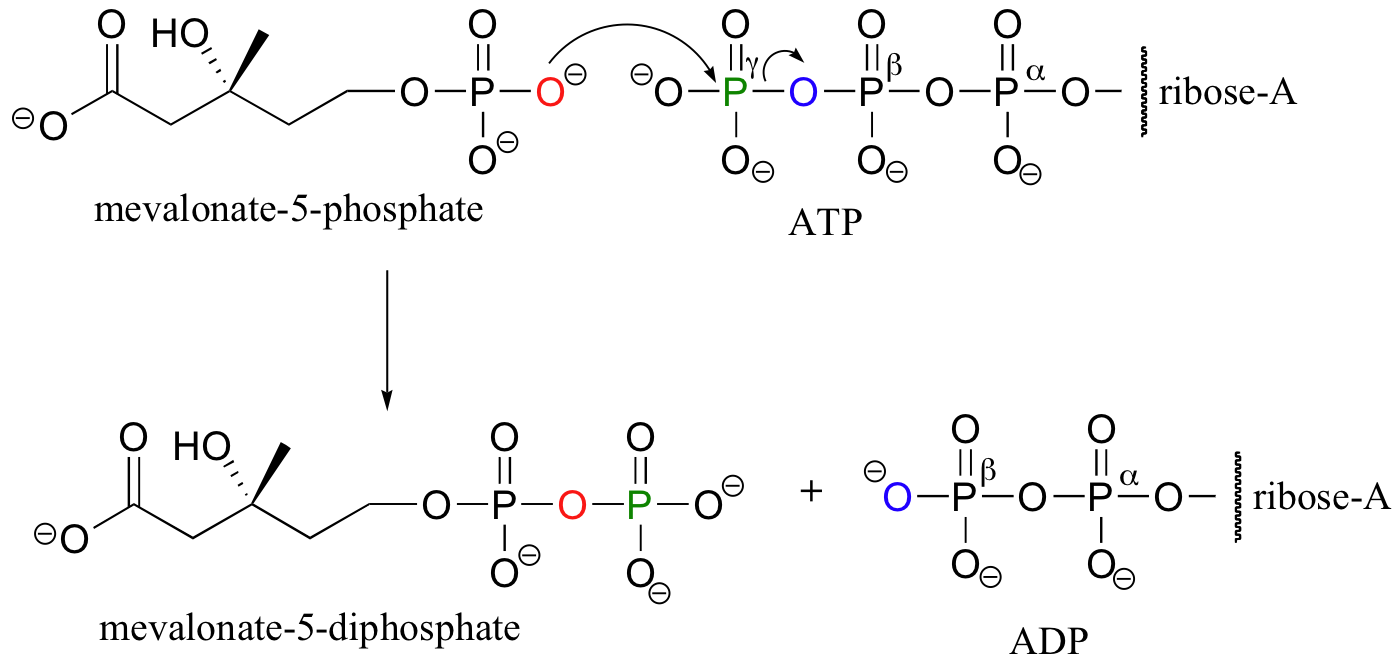
In the second phosphorylation reaction, the gamma phosphate of a second ATP molecule is transferred to an oxygen atom on the first phosphate, forming a new phosphate anhydride linkage.
Another example of a diphosphorylation reaction takes place in a single step, rather than sequentially. Phosphoribosyl diphosphate (PRPP) is a very important intermediate compound in the biosynthesis of nucleotides and some amino acids, and is the product of the diphosphorylation of ribose-5-phosphate.
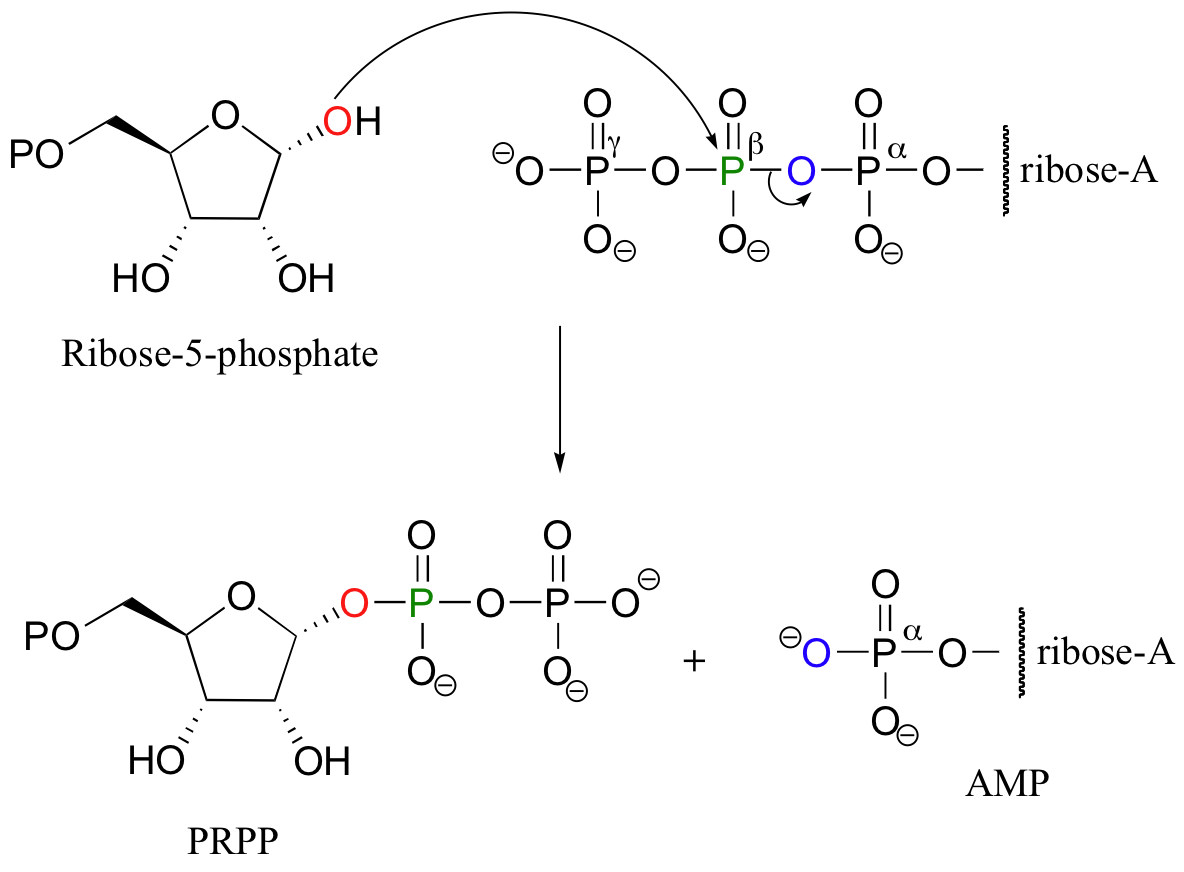
In this reaction, two phosphate groups (the beta and the gamma) are transferred together from ATP to a hydroxyl on ribose-5-phosphate. Notice that the beta phosphorus of ATP is the electrophile in this case, rather then the alpha phosphorus. Consequentially, the reaction results in the conversion on one ATP to one AMP, rather than two ATPs to two ADPs.
Are we getting more for our 'ATP money' in this one-step diphosphorylation? Not really - in order to convert the energy-poor AMP back up to the energy-rich ATP, the cell first has to transfer a phosphate from a second ATP molecule in a reaction catalyzed by an enzyme called adenylate kinase.
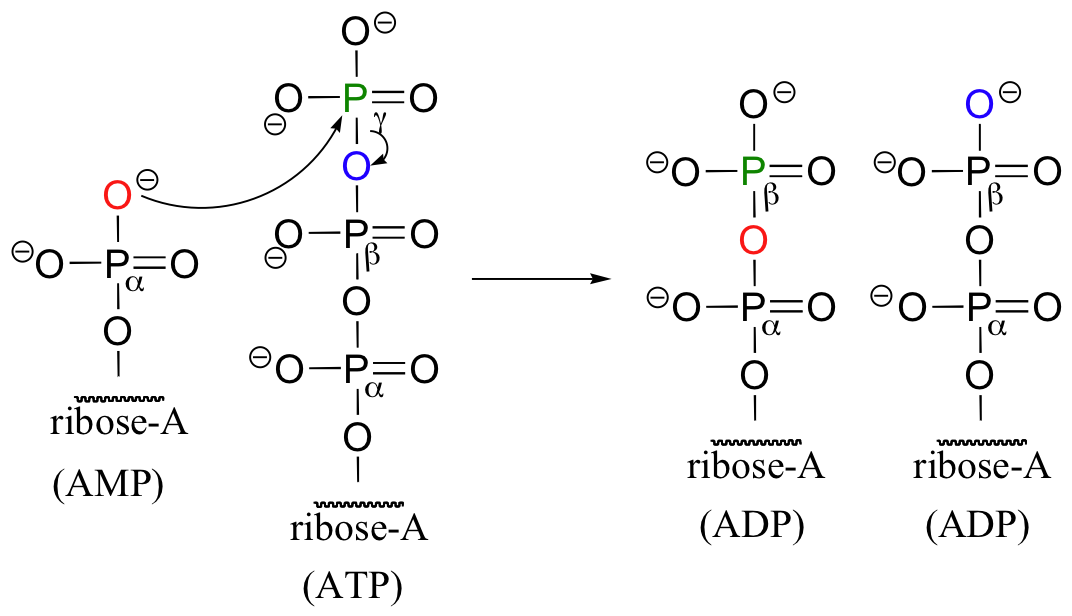
So in the end, the diphosphorylation reaction still costs the cell two ATP-to-ADP conversions.
It is worth noting that both of the diphosphate groups produced in these reactions end up as leaving groups in subsequent nucleophilic substitution reactions. Mevalonate diphosphate is eventually converted to isoprenyl diphosphate, the substrate for protein prenyltransferase (section 9.3). PRPP is the starting point for the biosynthesis of both pyrimidine (C and U/T) and purine (G, and A) nucleotides. The SN1 displacement of pyrophosphate in pyrimidine biosynthesis is shown below.

10.2D: Phosphorylation of carboxylates
Thus far we have seen alcohol oxygens and phosphate oxygens acting as nucleophilic accepting groups in phosphoryl transfer reactions. Consider next the first step of the reaction catalyzed by the enzyme glutamine synthase:
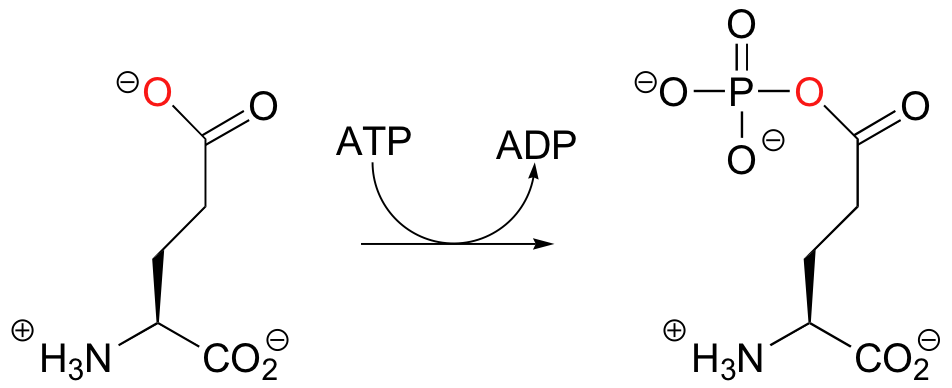
Once again in this reaction, the gamma-phosphate of ATP is transferred to an oxygen acceptor - however in this case the acceptor is a carboxylate oxygen, and the product is an acyl phosphate. As we shall see in chapter 12, acyl phosphates are commonly referred to as 'activated carboxylates', and are primed to undergo reactions called 'nucleophilic acyl substitutions'.
Example 10.5
10.2E: Generation of nucleotide phosphates
Activation of the side chain carboxylate of aspartate is somewhat different from the parallel activation of glutamate shown above. While the carboxylate group in glutamate accepts a simple phosphate group from ATP, the carboxylate in aspartate attacks the alpha-phosphate of ATP, displacing inorganic pyrophosphate and accepting an entire AMP group.
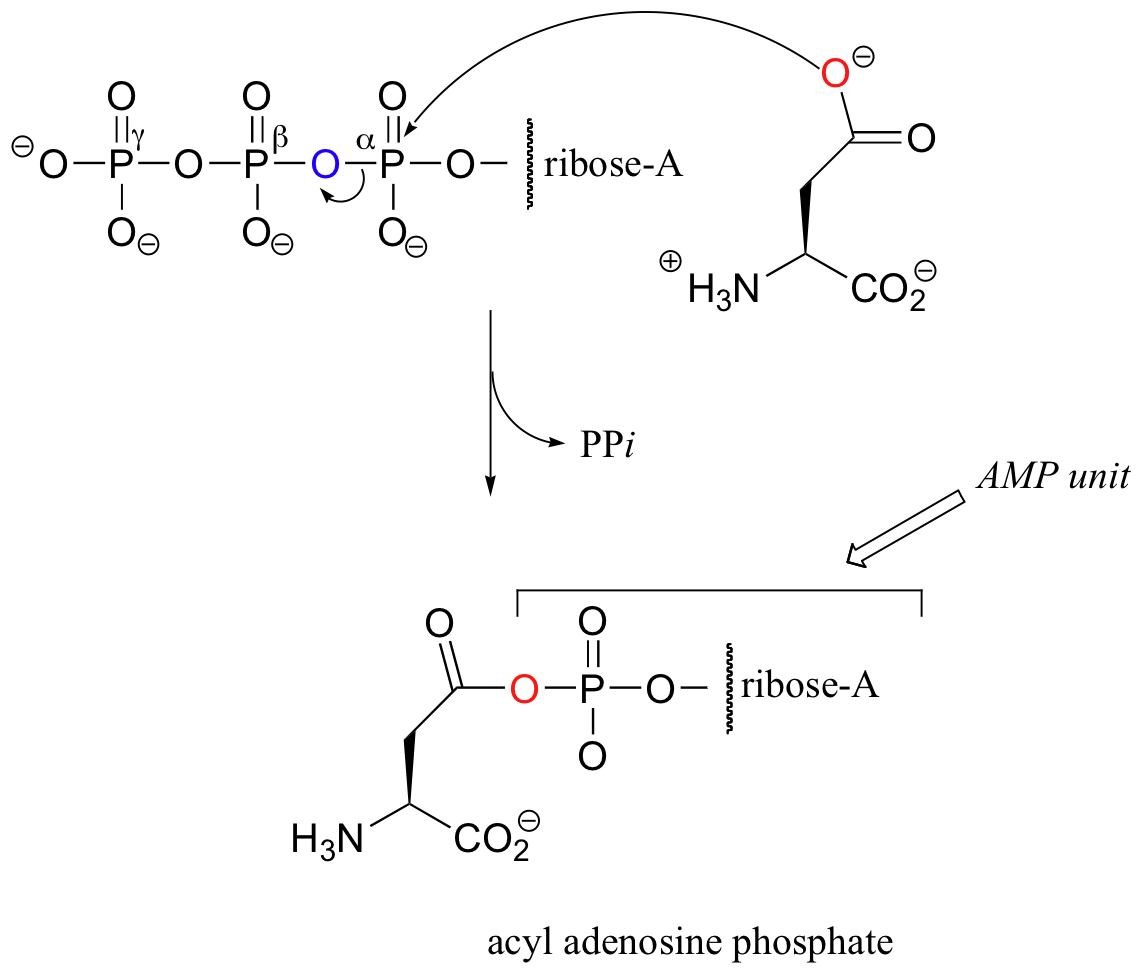
The resulting 'acyl adenosine phosphate', which is technically a phosphate diester, is another form of 'activated carboxylate' that we will learn more about in chapter 12.
For some interesting variations on the phosphoryl transfer reaction, consider the early steps of isoprenoid biosynthesis in bacteria (this is a completely different pathway than that mentioned in section 10.2C, which is operative in animals). In the first step, the oxygen of a monophosphate ester attacks the a phosphate of CTP (not ATP!) to expel inorganic pyrophosphate.

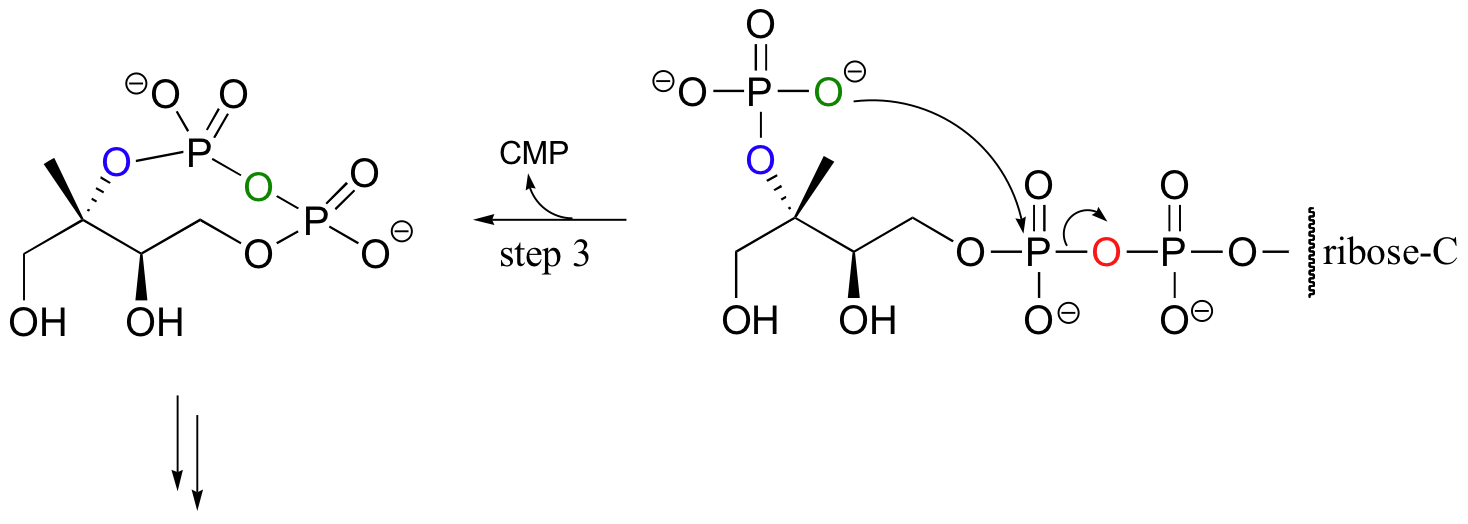

In step 2, a second hydroxyl group is phosphorylated in the normal way by an ATP-dependent kinase, and in step 3 that phosphate proceeds to attack the first of the two phosphates on the nucleotide diphosphate diester, expelling CMP and forming a cyclic diphosphate. Several more steps lead to the formation of isopentenyl diphosphate, the building block molecule for all isoprenoid compounds.
10.2F: Regeneration of ATP from ADP
Throughout this section we have seen reactions in which the energy contained in an ATP anhydride bond is 'spent', and ADP or AMP is formed as a result. In order to regenerate ATP, a phosphate group must be transferred to ADP, which is of course a thermodynamically uphill reaction requiring the input of energy from the breakdown of fuel molecules or, in the case of plants, from sunlight. By far the most important source of ATP regeneration is the enzyme ATP synthase, which catalyzes the direct condensation between inorganic phosphate and ADP.

Despite the apparent simplicity of the chemistry going on here, ATP synthase is an extremely large, complex, and fascinating enzyme, with multiple protein subunits and a intricate 'molecular motor' design. The reaction must be 'driven' uphill by using the energy from a proton gradient that is set up across the inner mitochondrial membrane. You will learn much more about this amazing biochemical machine if you take a course in biochemistry.
Two other reactions in the gycolytic (sugar breakdown) pathway also result in the generation of ATP from ADP, but these are minor physiological sources of ATP compared to ATP synthase. Phosphoglycerate kinase (named according to the reverse of the reaction shown below) transfers a phosphate from an acyl phosphate to ADP.
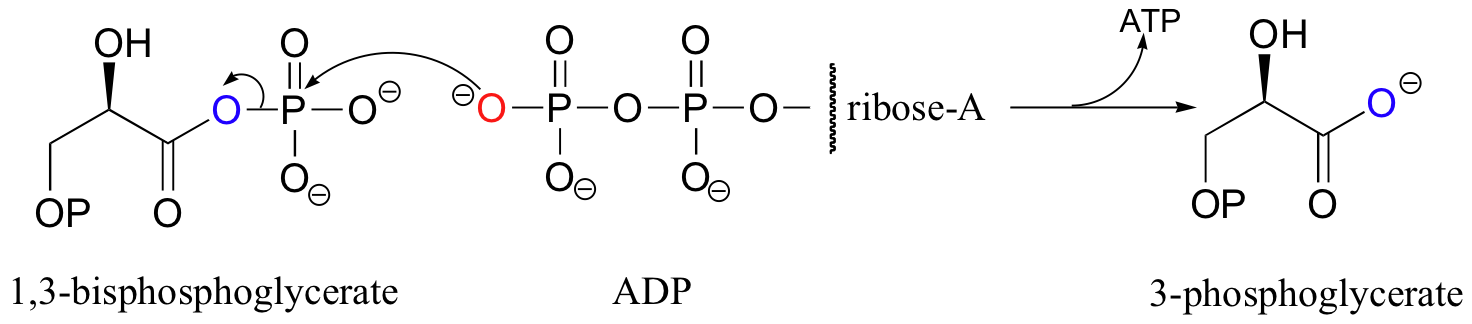
Note that here the leaving group is a carboxylate group.
Pyruvate kinase (again the name refers to the reverse reaction) catalyzes a less familiar-looking phosphate transfer.
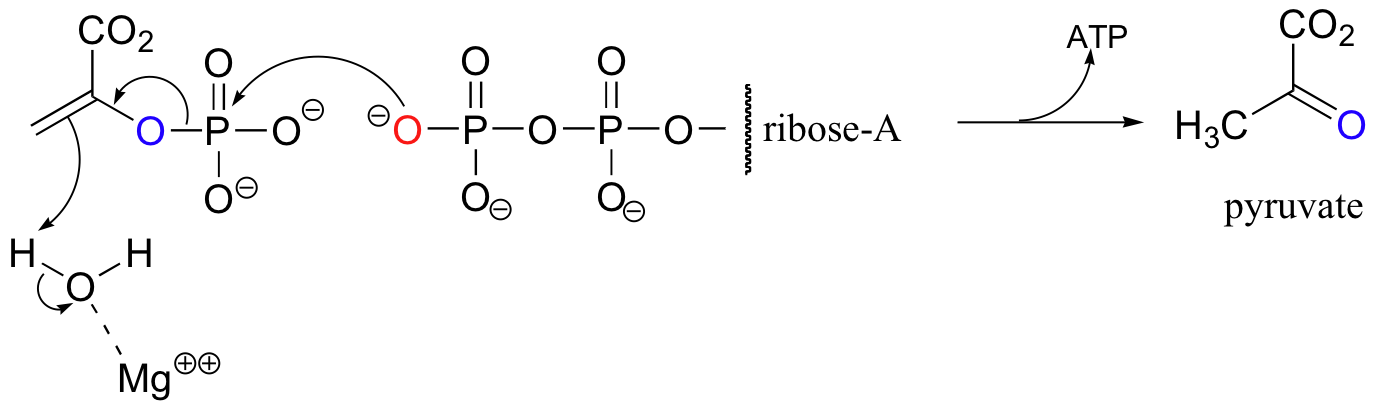
We will revisit this reaction in section 13.1A.