
9.2: Digestión de carbohidrato mediante glicosidasas - una reacción SN1
- Page ID
- 2378

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)A very important example of a nucleophilic substitution reaction that is thought to occur though a two-step, dissociative (SN1) mechanism is the hydrolytic breakdown of carbohydrates. In chapter 11 we will focus on these reactions and learn that the terms 'hemiacetal', 'acetal', 'hemiketal', and 'ketal' are used to describe the different patterns of bonding that we are seeing here - but for now, we will just look at the reaction from the standpoint of the SN1 mechanism that we learned about in chapter 8.
You probably know that carbohydrates, (also called oligosaccharides), are sugar polymers. Cellulose, the tough, fibrous material in plants, is one of the most abundant organic compounds found in nature. It is basically a long chain of glucose molecules linked together:
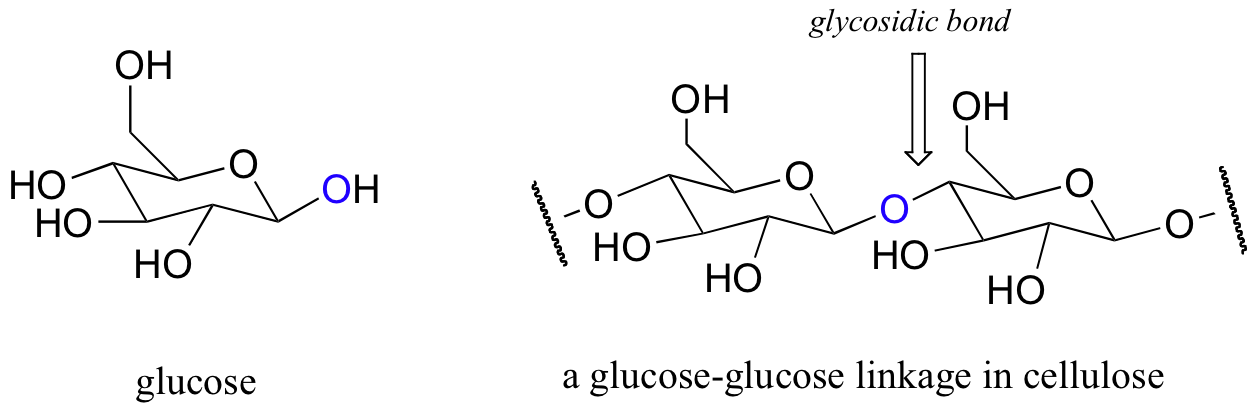
The bonds connecting individual sugar units in carbohydrate chains are called glycosidic bonds, and enzymes that break them are called glycosidases. Humans do not have glycosidases capable of breaking the glycosidic bonds in cellulose - and thus we cannot use cellulose a source of energy - but cellulose is nonetheless necessary to us as dietary fiber. Cows and other ruminants are able to derive energy from cellulose because they maintain bacteria in their digestive tract which possess the proper glycosidase enzymes.
Starch, the carbohydrate that we eat in bread and pasta, is also a long chain of glucose molecules. However, the glycosidic bonds linking the individual glucose units in starch have a different stereochemical configuration from those in cellulose, and humans do have the proper glycosidase enzymes to digest starch. We will discuss carbohydrates again in chapter 11, and you will learn many more details if you take a course in biochemistry.
We focus now on the chemistry by which glycosidic bonds are broken and formed. Enzymes called glycosidases catalyze these reactions. Consider for example the following reaction, catalyzed by a bacterial enzyme, in which the glycosidic bond between two glucose molecules in cellulose is cleaved.
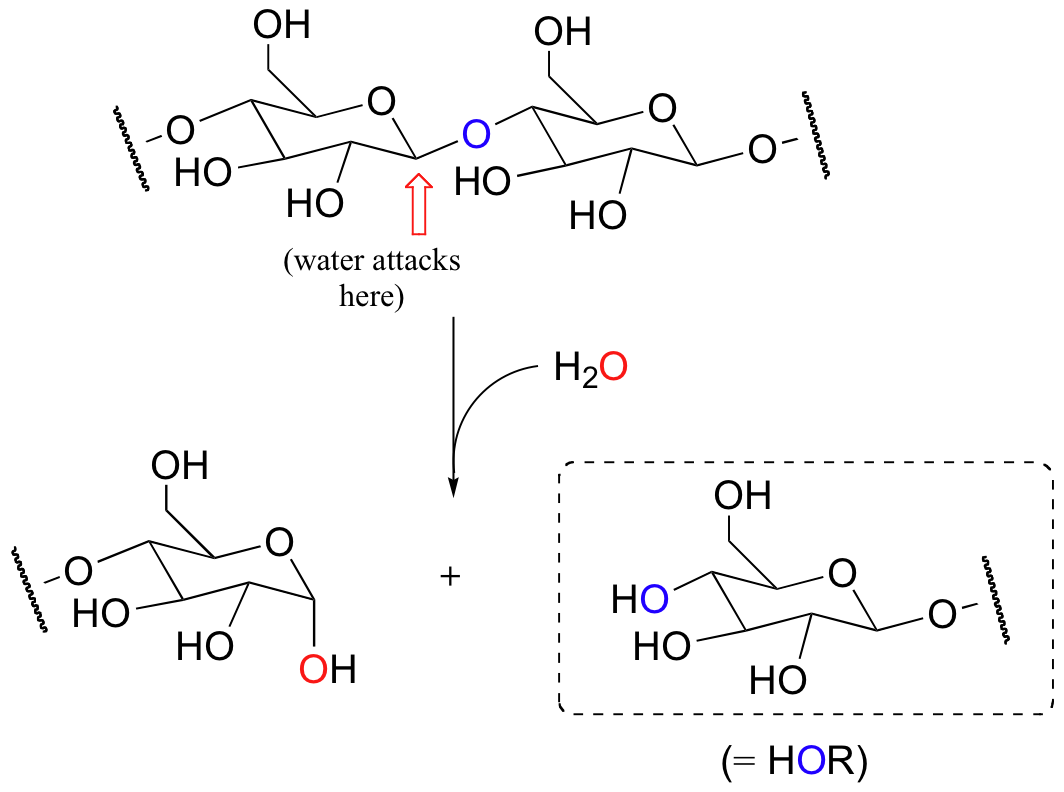
This is a hydrolysis reaction – recall that this term is used to describe any reaction where a bond is being broken by water. If you look carefully, you will recognize that this reaction is simply a nucleophilic substitution at the carbon indicated by the arrow: water is the nucleophile, and the leaving group is an alcohol, specifically the glucose molecule on the right side. In the next few figures, the leaving group will be referred to as 'HO-R' for simplicity.
(A quick note on hydrolysis reactions: we will see many more examples of hydrolysis reactions throughout our study of organic chemistry. In eukaryotic cells, many hydrolysis reactions occur in the acidic (pH ~4.5) environment of the lysosome, an organelle that specializes in breaking large molecules down into small ones.)
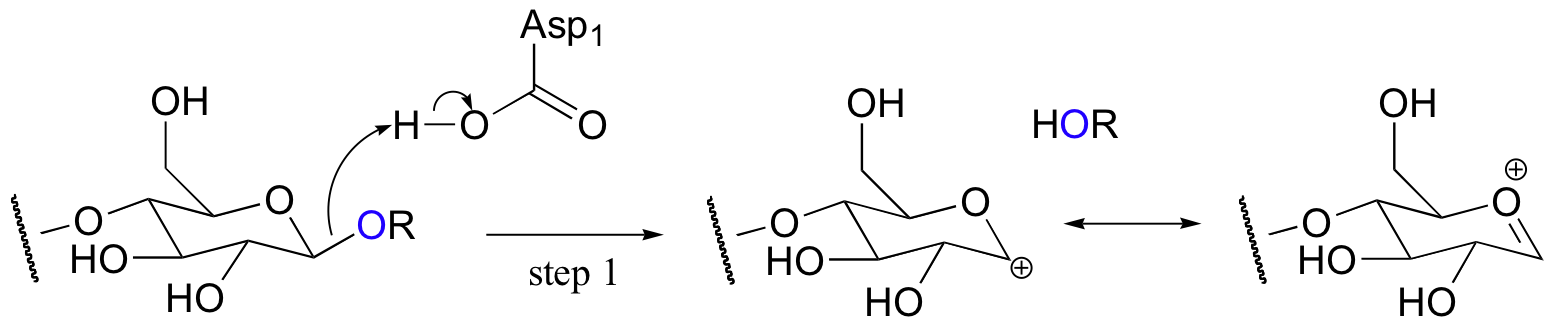
Evidence suggests that glycosidase reactions probably occur through an SN1 mechanism, implying the formation of a short-lived cationic intermediate. Here is the first, rate-determining step:
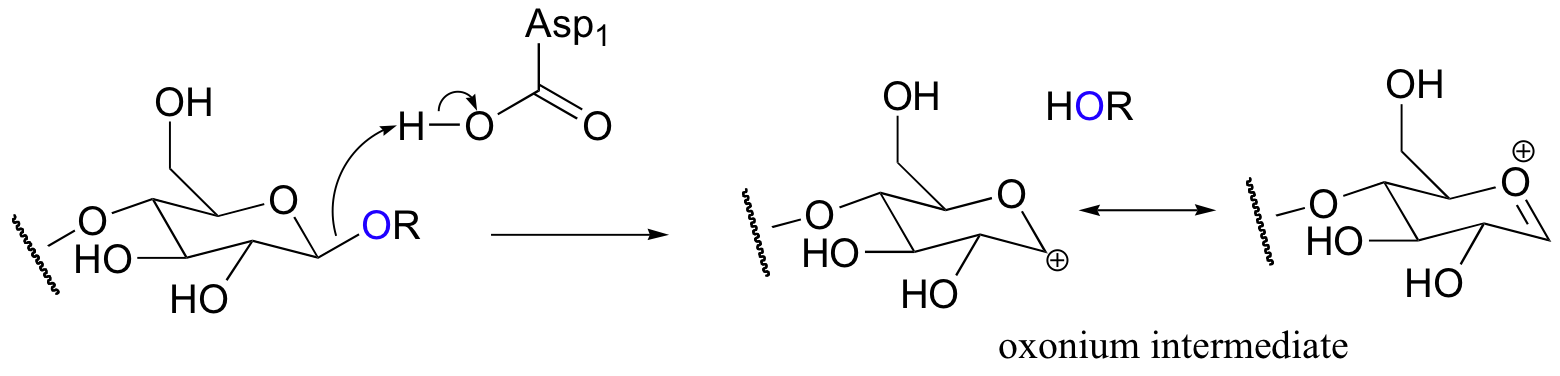
Notice that the positively charged carbon on the intermediate is adjacent to an oxygen. Recall from previous discussions (section 8.4B) that oxygen can act as a powerful electron donating group because of the resonance effect of its lone pairs. This is best illustrated by drawing a second resonance contributor, in which the positive charge is placed on the oxygen. This intermediate is generally referred to as an oxonium ion.
The active site of the enzyme has two aspartate residues, one positioned above the substrate (Asp1) and one below (Asp2). The leaving group is protonated by Asp1: protonation, as you recall, creates a better leaving group.
Here is the second step of the glycosidase mechanism:
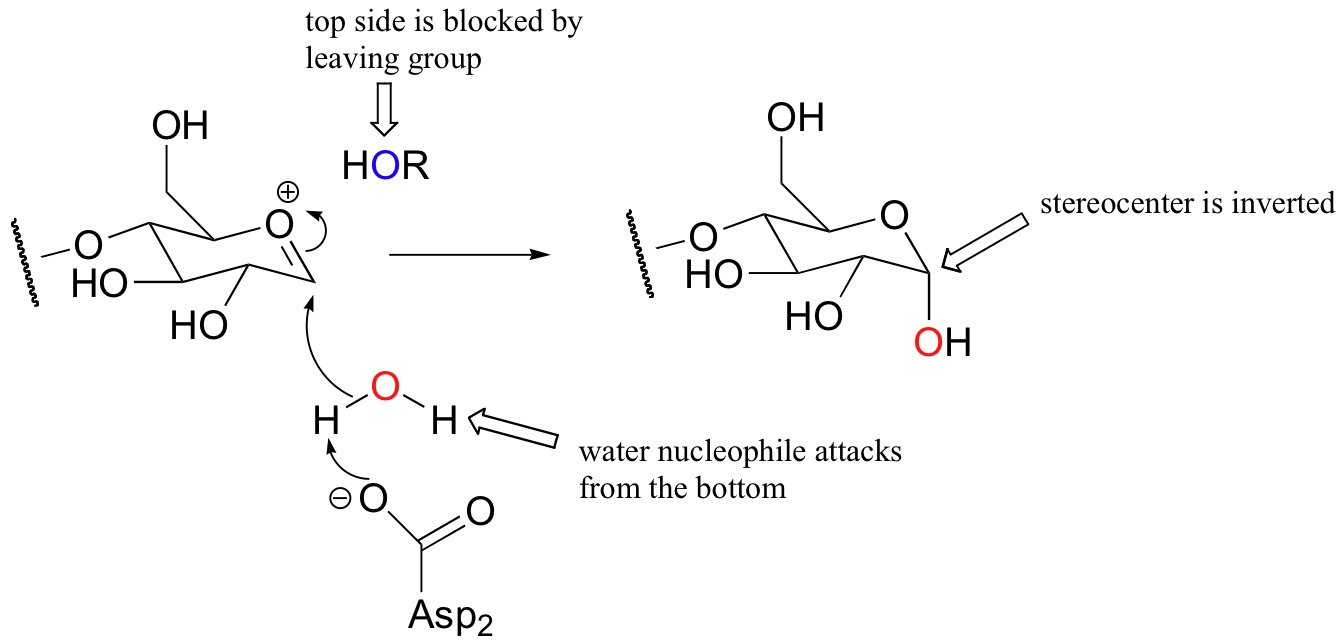
Notice that this substitution occurs with inversion of configuration:

This is because the leaving group remains bound in the active site after the formation of the oxonium ion intermediate, and blocks the bottom side of the electrophilic carbon from attack. The water nucleophile, as it attacks from the bottom side, is deprotonated by Asp2.
Because the reaction results in inversion of configuration, the enzyme is called an inverting glycosidase.
Researchers have also identified retaining glycosidases, which catalyze similar hydrolysis reactions except with retention of configuration:

The active site architectures of the inverting and retaining glycosidases are actually very similar: both have two aspartate residues positioned above and below the electrophilic carbon. The first step in the retaining mechanism is the same as in the inverting mechanism: formation of the oxonium intermediate.
In the retaining reaction, however, the bottom side aspartate (Asp2) acts as a nucleophile instead of as a base (step 2 below) , attacking the electrophilic carbon directly and forming what is referred to as a covalent intermediate (the enzyme is covalently attached to the terminal glucose molecule via the aspartate residue).
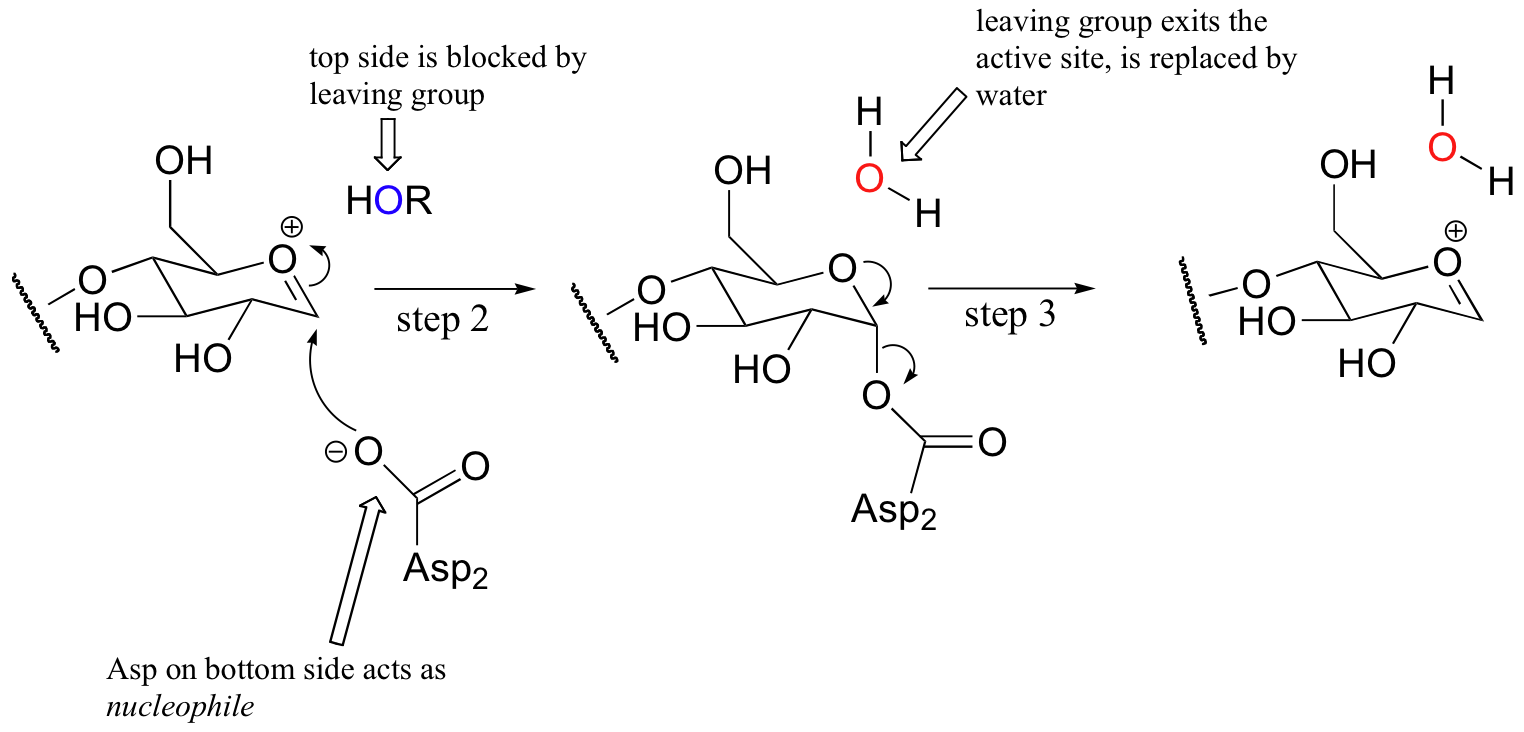
In the meantime, ROH (the other section of the celluose chain) diffuses away from the active site. The substrate-enzyme linkage then breaks (step 3), resulting in formation of the same oxonium ion intermediate as before. A nucleophilic water molecule then attacks from the top side, as Asp1 serves as a catalytic base (step 4).
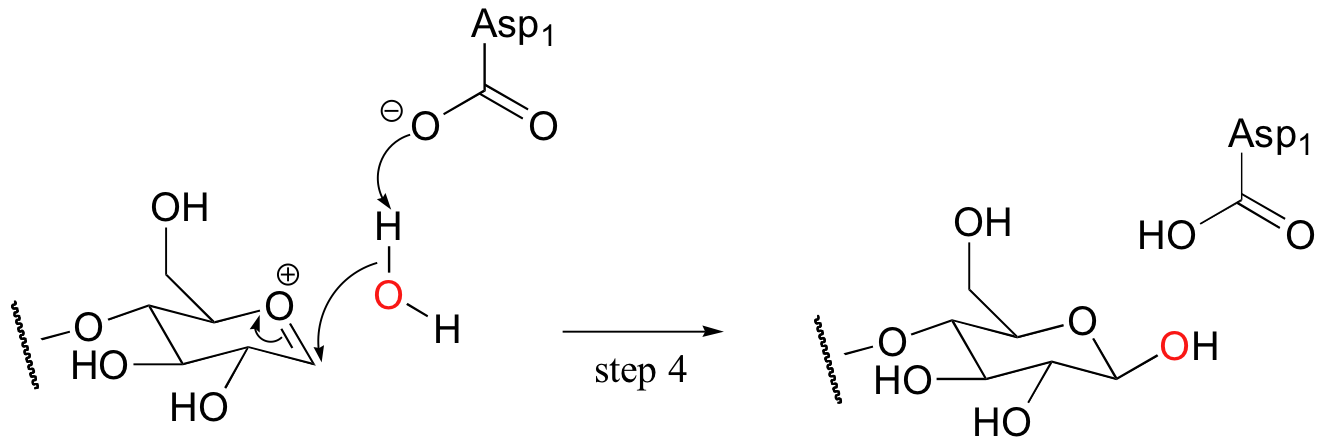
What has occurred here is known as a double displacement mechanism: in order to achieve substitution with retention of configuration, two nucleophilic substitutions have occurred in a row, each with inversion of stereochemistry. The overall result of two successive inversions at the same stereocenter is, of course, retention.
It is important to remember that the stereospecificity observed in the two examples of SN1 reactions above is due to the exquisite control that enzymes have over the reactionsthey catalyze. Nucleophiles and electrophiles are bound in precise locations so that nucleophilic attack can occur from one side and one side only. While concerted SN2 displacements take place specifically with inversion of configuration in both enzymatic and non-enzymatic situations, remember (section 8.2B) that nonenzymatic SN1 reactions generally result in racemization. Recall also the reasoning behind this: with no enzymatic control, the nucleophile can attack the planar, sp2-hybridized carbocation intermediate from either side with equal probability, resulting an a racemic mixture of both the R and S product.