
10.8: Músculo liso
- Page ID
- 122703

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de aprendizaje
- Describir un cuerpo denso
- Explicar cómo funciona el músculo liso con los órganos internos y los conductos a través del cuerpo
- Explicar cómo los músculos lisos difieren de los músculos esqueléticos y cardíacos
- Explicar la diferencia entre músculo liso de una sola unidad y multiunidad
El músculo liso (llamado así porque las células no tienen estrías) está presente en las paredes de órganos huecos como la vejiga urinaria, el útero, el estómago, los intestinos y en las paredes de los conductos, como las arterias y venas del sistema circulatorio, y los tractos de las vías respiratorias, urinarias y reproductivas sistemas (Figura\(\PageIndex{1}\) .a, b). El músculo liso también está presente en los ojos, donde funciona para cambiar el tamaño del iris y alterar la forma del cristalino; y en la piel donde hace que el cabello se ponga erecto en respuesta a la temperatura fría o al miedo.
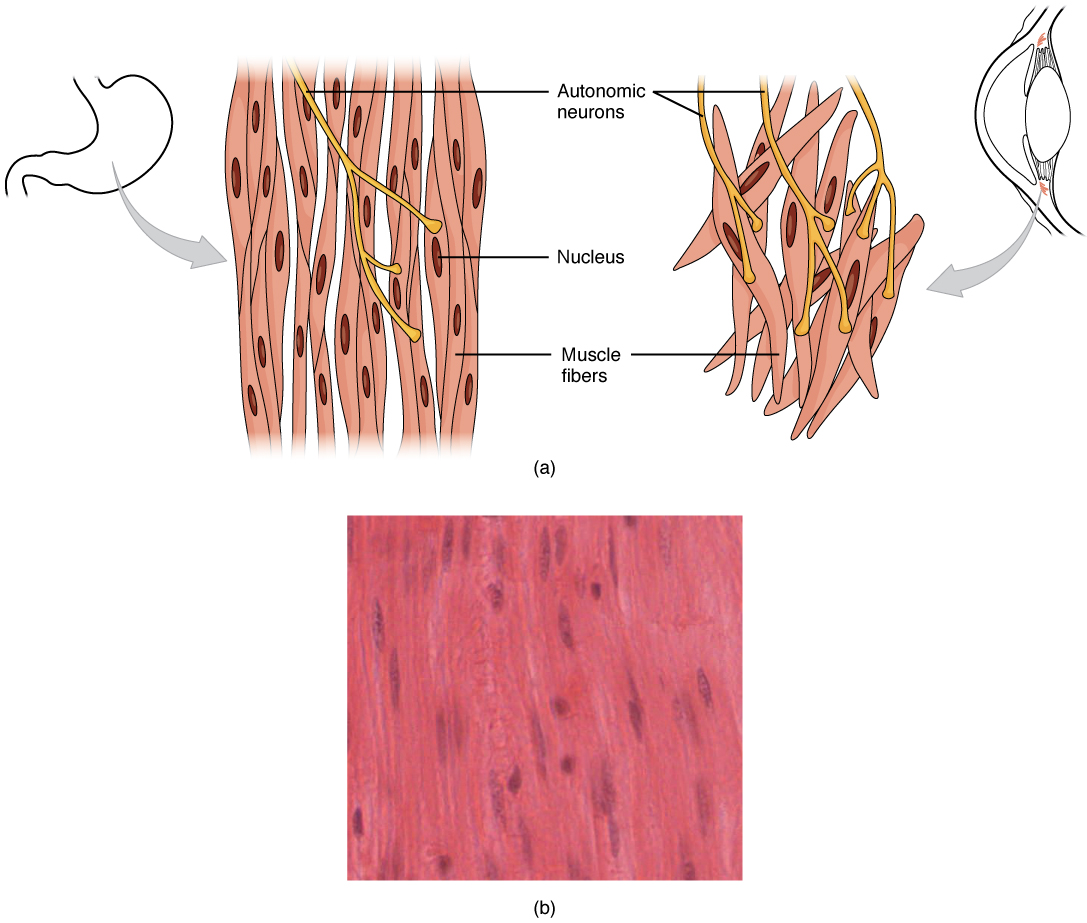

Las fibras musculares lisas tienen forma de huso (anchas en el medio y cónicas en ambos extremos, algo así como una pelota de fútbol) y tienen un solo núcleo; van de aproximadamente 30 a 200 μ m (miles de veces más cortas que las fibras del músculo esquelético), y producen su propio tejido conectivo, el endomisio. Aunque no tienen estriaciones y sarcómeros, las fibras musculares lisas sí tienen proteínas contráctiles de actina y miosina, y filamentos gruesos y delgados. Estos finos filamentos están anclados por cuerpos densos. Un cuerpo denso es análogo a los discos Z de las fibras del músculo esquelético y cardíaco y se sujeta al sarcolema. Los iones de calcio son suministrados por la SR en las fibras y por secuestro del fluido extracelular a través de indentaciones de membrana llamadas calveoli.
Debido a que las células del músculo liso no contienen troponina, la formación de puentes cruzados no está regulada por el complejo troponina-tropomiosina sino por la proteína reguladora calmodulina. En una fibra de músculo liso, los iones externos de Ca ++ que pasan a través de canales de calcio abiertos en el sarcolema, y el Ca ++ adicional liberado de SR, se unen a la calmodulina. El complejo Ca ++ -calmodulina activa entonces una enzima llamada miosina (cadena ligera) quinasa, que, a su vez, activa las cabezas de miosina fosforilándolas (convirtiendo ATP en ADP y P i, con el P i unido a la cabeza). Las cabezas se pueden unir a los sitios de unión de actina y tirar de los filamentos delgados. Los filamentos delgados también están anclados a los cuerpos densos; las estructuras invertidas en la membrana interna del sarcolema (en uniones adherentes) que también tienen filamentos intermedios en forma de cordón unidos a ellos. Cuando los filamentos delgados se deslizan más allá de los filamentos gruesos, tiran de los cuerpos densos, estructuras atadas al sarcolema, que luego tiran de las redes de filamentos intermedios a lo largo del sarcoplasma. Esta disposición hace que toda la fibra muscular se contraiga de manera que los extremos se estiren hacia el centro, haciendo que la sección media se abombe en un movimiento sacacorchos (Figura\(\PageIndex{2}\)).

Although smooth muscle contraction relies on the presence of Ca++ ions, smooth muscle fibers have a much smaller diameter than skeletal muscle cells. T-tubules are not required to reach the interior of the cell and therefore not necessary to transmit an action potential deep into the fiber. Smooth muscle fibers have a limited calcium-storing SR but have calcium channels in the sarcolemma (similar to cardiac muscle fibers) that open during the action potential along the sarcolemma. The influx of extracellular Ca++ ions, which diffuse into the sarcoplasm to reach the calmodulin, accounts for most of the Ca++ that triggers contraction of a smooth muscle cell.
Muscle contraction continues until ATP-dependent calcium pumps actively transport Ca++ ions back into the SR and out of the cell. However, a low concentration of calcium remains in the sarcoplasm to maintain muscle tone. This remaining calcium keeps the muscle slightly contracted, which is important in certain tracts and around blood vessels.
Because most smooth muscles must function for long periods without rest, their power output is relatively low, but contractions can continue without using large amounts of energy. Some smooth muscle can also maintain contractions even as Ca++ is removed and myosin kinase is inactivated/dephosphorylated. This can happen as a subset of cross-bridges between myosin heads and actin, called latch-bridges, keep the thick and thin filaments linked together for a prolonged period, and without the need for ATP. This allows for the maintaining of muscle “tone” in smooth muscle that lines arterioles and other visceral organs with very little energy expenditure.
Smooth muscle is not under voluntary control; thus, it is called involuntary muscle. The triggers for smooth muscle contraction include hormones, neural stimulation by the ANS, and local factors. In certain locations, such as the walls of visceral organs, stretching the muscle can trigger its contraction (the stretch-relaxation response).
Axons of neurons in the ANS do not form the highly organized NMJs with smooth muscle, as seen between motor neurons and skeletal muscle fibers. Instead, there is a series of neurotransmitter-filled bulges called varicosities as an axon courses through smooth muscle, loosely forming motor units (Figure \(\PageIndex{3}\)). A varicosity releases neurotransmitters into the synaptic cleft. Also, visceral muscle in the walls of the hollow organs (except the heart) contains pacesetter cells. A pacesetter cell can spontaneously trigger action potentials and contractions in the muscle.

Smooth muscle is organized in two ways: as single-unit smooth muscle, which is much more common; and as multiunit smooth muscle. The two types have different locations in the body and have different characteristics. Single-unit muscle has its muscle fibers joined by gap junctions so that the muscle contracts as a single unit. This type of smooth muscle is found in the walls of all visceral organs except the heart (which has cardiac muscle in its walls), and so it is commonly called visceral muscle. Because the muscle fibers are not constrained by the organization and stretchability limits of sarcomeres, visceral smooth muscle has a stress-relaxation response. This means that as the muscle of a hollow organ is stretched when it fills, the mechanical stress of the stretching will trigger contraction, but this is immediately followed by relaxation so that the organ does not empty its contents prematurely. This is important for hollow organs, such as the stomach or urinary bladder, which continuously expand as they fill. The smooth muscle around these organs also can maintain a muscle tone when the organ empties and shrinks, a feature that prevents “flabbiness” in the empty organ. In general, visceral smooth muscle produces slow, steady contractions that allow substances, such as food in the digestive tract, to move through the body.
Multiunit smooth muscle cells rarely possess gap junctions, and thus are not electrically coupled. As a result, contraction does not spread from one cell to the next, but is instead confined to the cell that was originally stimulated. Stimuli for multiunit smooth muscles come from autonomic nerves or hormones but not from stretching. This type of tissue is found around large blood vessels, in the respiratory airways, and in the eyes.
Hyperplasia in Smooth Muscle
Similar to skeletal and cardiac muscle cells, smooth muscle can undergo hypertrophy to increase in size. Unlike other muscle, smooth muscle can also divide to produce more cells, a process called hyperplasia. This can most evidently be observed in the uterus at puberty, which responds to increased estrogen levels by producing more uterine smooth muscle fibers, and greatly increases the size of the myometrium.
Sections Summary
Smooth muscle is found throughout the body around various organs and tracts. Smooth muscle cells have a single nucleus, and are spindle-shaped. Smooth muscle cells can undergo hyperplasia, mitotically dividing to produce new cells. The smooth cells are nonstriated, but their sarcoplasm is filled with actin and myosin, along with dense bodies in the sarcolemma to anchor the thin filaments and a network of intermediate filaments involved in pulling the sarcolemma toward the fiber’s middle, shortening it in the process. Ca++ ions trigger contraction when they are released from SR and enter through opened voltage-gated calcium channels. Smooth muscle contraction is initiated when the Ca++ binds to intracellular calmodulin, which then activates an enzyme called myosin kinase that phosphorylates myosin heads so they can form the cross-bridges with actin and then pull on the thin filaments. Smooth muscle can be stimulated by pacesetter cells, by the autonomic nervous system, by hormones, spontaneously, or by stretching. The fibers in some smooth muscle have latch-bridges, cross-bridges that cycle slowly without the need for ATP; these muscles can maintain low-level contractions for long periods. Single-unit smooth muscle tissue contains gap junctions to synchronize membrane depolarization and contractions so that the muscle contracts as a single unit. Single-unit smooth muscle in the walls of the viscera, called visceral muscle, has a stress-relaxation response that permits muscle to stretch, contract, and relax as the organ expands. Multiunit smooth muscle cells do not possess gap junctions, and contraction does not spread from one cell to the next.
Multiple Choice
Q. Smooth muscles differ from skeletal and cardiac muscles in that they ________.
A. lack myofibrils
B. are under voluntary control
C. lack myosin
D. lack actin
Answer: A
Q. Which of the following statements describes smooth muscle cells?
A. They are resistant to fatigue.
B. They have a rapid onset of contractions.
C. They cannot exhibit tetanus.
D. They primarily use anaerobic metabolism.
Answer: A
Free Response
Q. Why can smooth muscles contract over a wider range of resting lengths than skeletal and cardiac muscle?
A. Smooth muscles can contract over a wider range of resting lengths because the actin and myosin filaments in smooth muscle are not as rigidly organized as those in skeletal and cardiac muscle.
Q. Describe the differences between single-unit smooth muscle and multiunit smooth muscle.
A. Single-unit smooth muscle is found in the walls of hollow organs; multiunit smooth muscle is found in airways to the lungs and large arteries. Single-unit smooth muscle cells contract synchronously, they are coupled by gap junctions, and they exhibit spontaneous action potential. Multiunit smooth cells lack gap junctions, and their contractions are not synchronous.
Glossary
- calmodulin
- regulatory protein that facilitates contraction in smooth muscles
- dense body
- sarcoplasmic structure that attaches to the sarcolemma and shortens the muscle as thin filaments slide past thick filaments
- hyperplasia
- process in which one cell splits to produce new cells
- latch-bridges
- subset of a cross-bridge in which actin and myosin remain locked together
- pacesetter cell
- cell that triggers action potentials in smooth muscle
- stress-relaxation response
- relaxation of smooth muscle tissue after being stretched
- varicosity
- enlargement of neurons that release neurotransmitters into synaptic clefts
- visceral muscle
- smooth muscle found in the walls of visceral organs