
2.8: Epigenética
- Page ID
- 124518

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Hemos visto cómo la acción de los neurotransmisores puede alterar la transcripción y traducción de genes a través de la unión a receptores acoplados a proteínas G. La efectividad de la cascada de señalización en la síntesis de nuevas proteínas depende de algunos factores específicos del ADN. Este capítulo cubrirá brevemente cómo se transcriben los genes y luego cómo los cambios moleculares no secuenciales en el ADN pueden afectar las tasas de transcripción.
Dogma Central
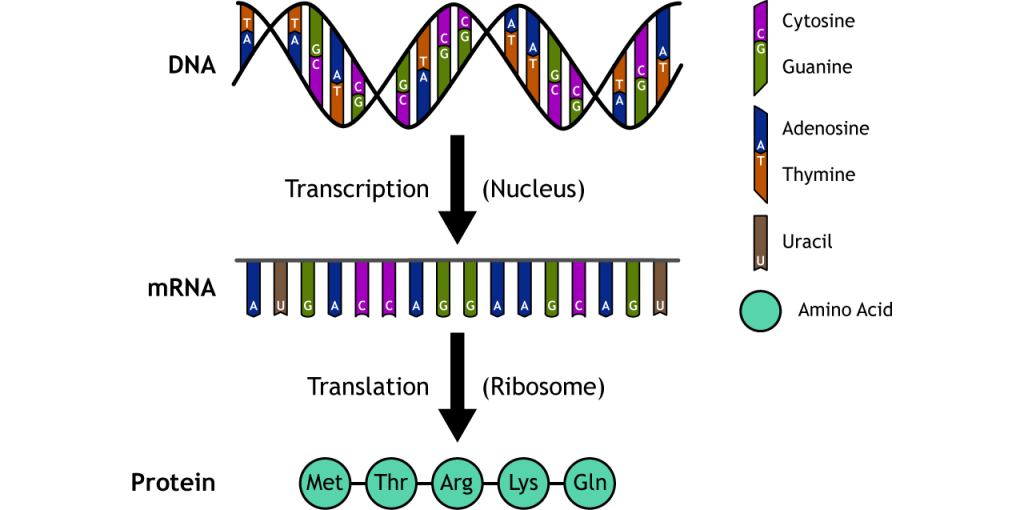
ADN a ARN a proteína. El dogma central de la genética. Puede parecer simple, pero deben darse muchos pasos complejos para que el proceso sea exitoso.
ADN
El ADN bicatenario (ácido desoxirribonucleico) está compuesto por cuatro bases nucleotídicas: adenosina (A), timina (T), guanina (G) y citosina (C). La adenosina y la timina forman pares de bases, mientras que la guanina y la citosina forman pares. Los pares hacen que los dos hilos se enrollen uno alrededor del otro y formen una doble hélice.
RNA
El ARN mensajero monocatenario (ácido ribonucleico) se crea a partir de la secuencia de ADN mediante emparejamiento de bases complementarias. Al igual que el ADN, hay cuatro bases, pero en el ARN la base de timina es reemplazada por uracilo (U). El ARN mensajero (ARNm) sale del núcleo e interactúa con los ribosomas para sintetizar proteínas en un proceso llamado traducción. Los ribosomas emparejan aminoácidos con secuencias específicas de tres bases llamadas codones. Por ejemplo, la secuencia de codones AUG es el codón de inicio y codifica metionina. Los ribosomas bajarán por el ARNm para encontrar el codón de inicio de la proteína y comenzar la traducción allí, agregando un nuevo aminoácido para cada codón hasta que se alcance un codón de parada.
Proteína
Las proteínas son sintetizadas por la unión de aminoácidos entre sí por los ribosomas. Hay 20 aminoácidos que están codificados cada uno por una o más secuencias de codones de ARNm.
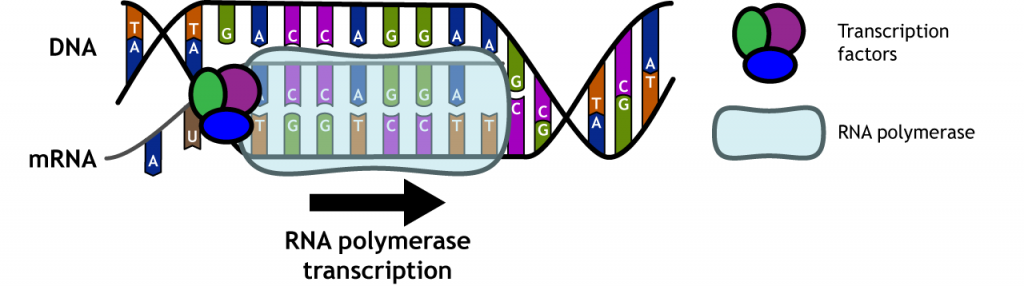
Transcripción de genes
En el núcleo, proteínas llamadas factores de transcripción y una ARN polimerasa se adhieren al ADN. El ADN se desenrolla, las proteínas se unen y se sintetiza una cadena de ARNm usando el ADN como molde. El ARNm es una secuencia complementaria a la cadena de ADN que se transcribe.
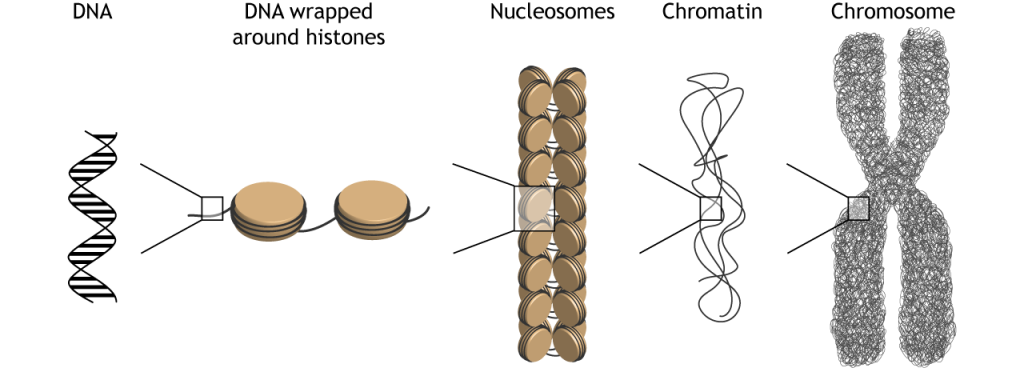
Envasado de ADN
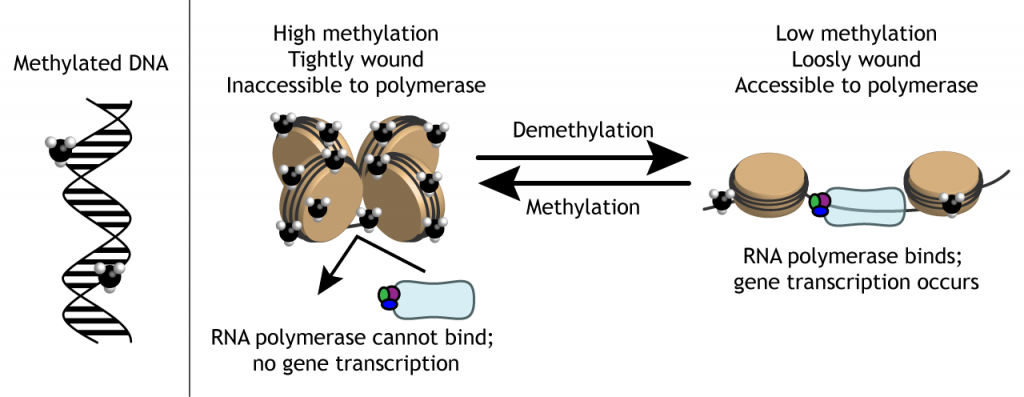
Sin embargo, el ADN no siempre es accesible para esas proteínas de transcripción. Hay tanto ADN en cada célula, que para ahorrar espacio, está altamente condensado en el núcleo. La doble hélice se envuelve alrededor de proteínas llamadas histonas. Luego, las histonas se envuelven en hebras de nucleosomas. Los nucleosomas se compactan en estructuras más densas llamadas cromatina. Finalmente, la cromatina se condensa más y crea cromosomas.
Para que se produzca la transcripción génica, las cadenas de ADN deben desenrollarse de los cuerpos de histonas para ser accesibles a la maquinaria transcripcional.
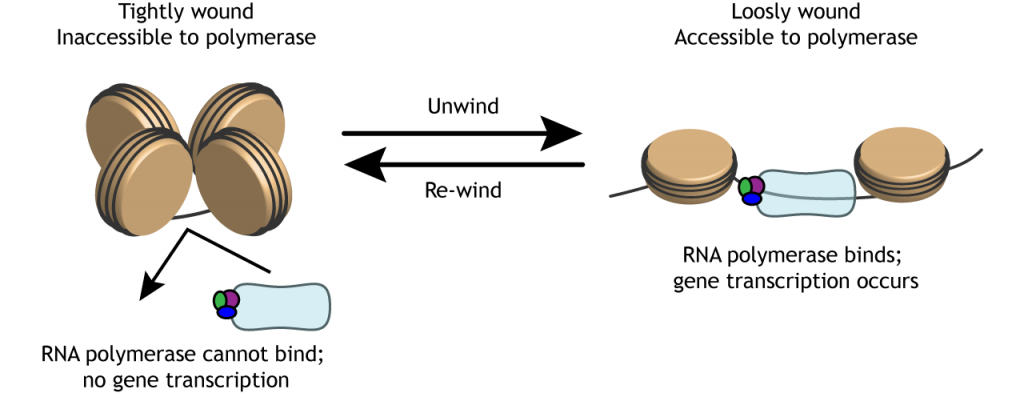
Epigenética
Moléculas como los grupos metilo pueden unirse al ADN o a las histonas. Estas etiquetas epigenéticas pueden afectar la fuerza con la que se enrolla el ADN alrededor de las histonas. Dado que la expresión génica puede alterarse modificando la facilidad con la que las histonas se desenrollan y cuán accesibles son las cadenas de ADN, las etiquetas epigenéticas pueden tener un efecto indirecto sobre la transcripción génica.
Los grupos metilo dificultan que la polimerasa acceda al ADN al mantener el ADN enrollado alrededor de las histonas, reduciendo la transcripción. Cuando se eliminan los grupos metilo, llamada desmetilación (que no debe confundirse con la dimetilación, la adición de dos grupos metilo), la expresión génica puede aumentar debido a que el ADN se desenrolla y es accesible a la maquinaria transcripcional.
El epigenoma es flexible
La secuencia de ADN de un individuo es fija (excluyendo las mutaciones que ocurren por daños o errores en la replicación celular), pero el epigenoma es flexible y puede cambiar a lo largo de la vida. Las experiencias de vida de un individuo, especialmente durante el desarrollo u otros períodos críticos, son capaces de alterar el epigenoma.
Algunas experiencias aumentarán la metilación, a veces solo para ciertos genes, a veces en todo el genoma, mientras que otras experiencias la disminuirán. Por ejemplo, el estrés temprano en la vida puede aumentar la cantidad de metilación que se encuentra en el gen que codifica para el receptor que es activado por las hormonas del estrés. El aumento de la metilación conduce a una transcripción reducida que tiene efectos aguas abajo en el bucle de retroalimentación negativa en la respuesta al estrés. Los científicos están empezando a darse cuenta de lo importante que es el epigenoma en la regulación de nuestro cerebro y comportamiento.
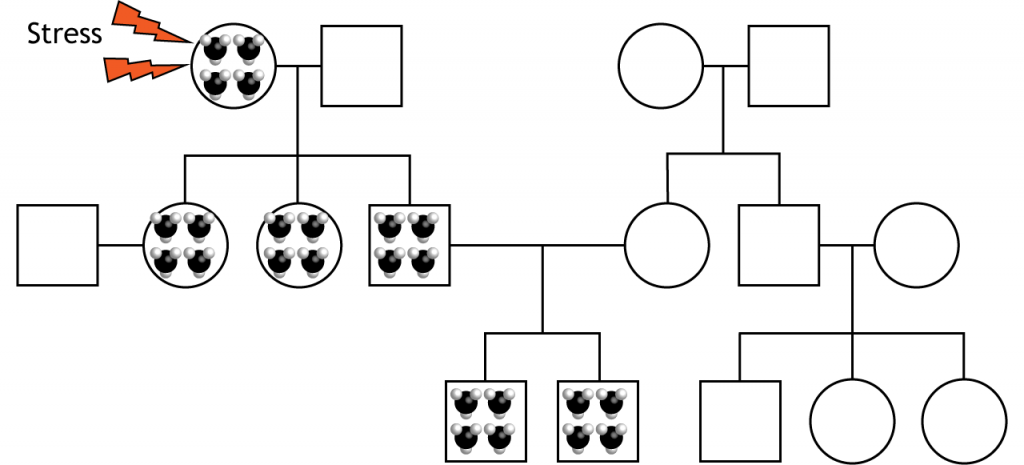
Epigenoma heredado
Adicionalmente, las modificaciones epigenéticas son heredables. Investigaciones recientes comienzan a mostrar que las experiencias de madres, padres e incluso abuelos pueden tener efectos transgeneracionales. Y estos efectos, que alguna vez se pensaba que sólo se heredaban del lado materno, ahora se ha demostrado que también son heredados paternalmente. Esto significa que un animal que tuvo estrés temprano en la vida puede haber aumentado la metilación y cambios en la transcripción génica que luego se transmite por generaciones incluso si la descendencia no experimenta los mismos factores estresantes.
Claves para llevar
- El ADN está altamente condensado en el núcleo
- El ADN debe desenrollarse para que se lleve a cabo la transcripción
- Las modificaciones epigenéticas pueden alterar la facilidad con la que se puede desenrollar el ADN
- Las modificaciones epigenéticas se pueden heredar
Versión en video de la lección

Un elemento de YouTube ha sido excluido de esta versión del texto. Puedes verlo en línea aquí: https://openbooks.lib.msu.edu/neuroscience/?p=609