
3.1: Transporte de gas
- Page ID
- 121074
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Objetivos de aprendizaje
- Explicar cómo se transportan el oxígeno y el dióxido de carbono en sangre o medio líquido equivalente.
La otra actividad importante en los pulmones es el proceso de respiración, el proceso de intercambio de gases. La función de la respiración es proporcionar oxígeno para su uso por las células del cuerpo durante la respiración celular y eliminar el dióxido de carbono, un producto de desecho de la respiración celular, del cuerpo. Para que se produzca el intercambio de oxígeno y dióxido de carbono, ambos gases deben ser transportados entre los sitios de respiración externa e interna. Aunque el dióxido de carbono es más soluble que el oxígeno en la sangre, ambos gases requieren de un sistema de transporte especializado para que la mayoría de las moléculas de gas se muevan entre los pulmones y otros tejidos.
Transporte de Oxígeno en la Sangre
A pesar de que el oxígeno se transporta a través de la sangre, es posible que recuerde que el oxígeno no es muy soluble en líquidos. Una pequeña cantidad de oxígeno sí se disuelve en la sangre y se transporta en el torrente sanguíneo, pero es sólo alrededor del 1.5% de la cantidad total. La mayoría de las moléculas de oxígeno son transportadas de los pulmones a los tejidos del cuerpo por un sistema de transporte especializado, que se basa en el eritrocito, el glóbulo rojo. Los eritrocitos contienen una metaloproteína, la hemoglobina, que sirve para unir moléculas de oxígeno al eritrocito (Figura 3.1). El hemo es la porción de hemoglobina que contiene hierro, y es el hemo el que se une al oxígeno. Una molécula de hemoglobina contiene moléculas Heme que contienen hierro, y debido a esto, cada molécula de hemoglobina es capaz de transportar hasta cuatro moléculas de oxígeno. A medida que el oxígeno se difunde a través de la membrana respiratoria desde el alvéolo hasta el capilar, también se difunde hacia el glóbulo rojo y se une a la hemoglobina. La siguiente reacción química reversible describe la producción del producto final, oxihemoglobina (\(\ce{Hb–O2}\)), que se forma cuando el oxígeno se une a la hemoglobina. La oxihemoglobina es una molécula de color rojo brillante que contribuye al color rojo brillante de la sangre oxigenada.
\[\ce{Hb + O2 <=> Hb − O2Hb + O2 <=> Hb − O2}\]
En esta fórmula, Hb representa hemoglobina reducida, es decir, hemoglobina que no tiene oxígeno unido a ella. Existen múltiples factores involucrados en la facilidad con la que el hemo se une y se disocia del oxígeno, lo que se discutirá en las secciones siguientes.
Eritrocitos y Hemoglobina
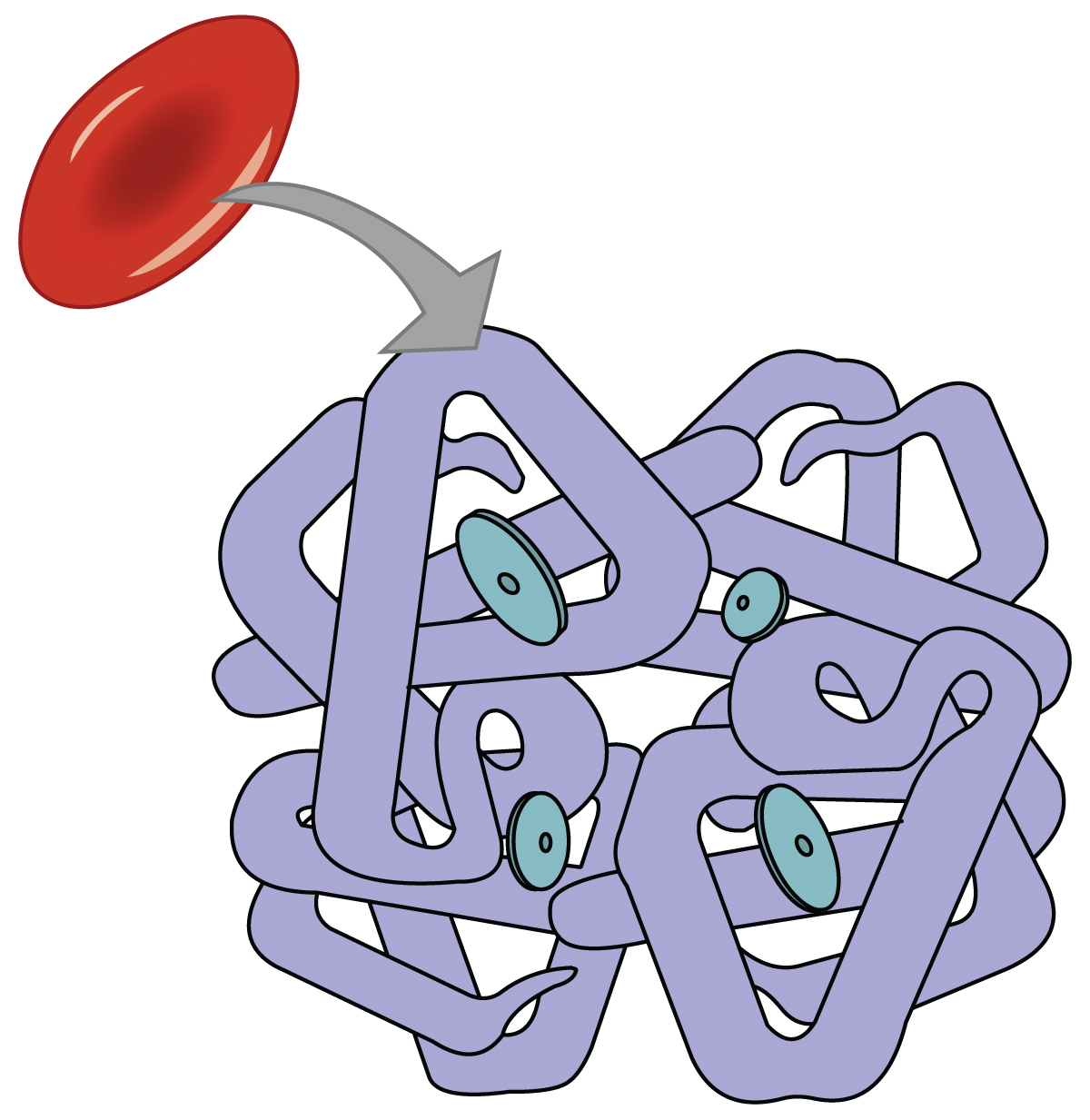
Figura 3.1. La hemoglobina consta de cuatro subunidades, cada una de las cuales contiene una molécula de hierro.
Función de la hemoglobina
La hemoglobina está compuesta por subunidades, una estructura proteica que se denomina estructura cuaternaria. Cada una de las cuatro subunidades que componen la hemoglobina está dispuesta en forma de anillo, con un átomo de hierro unido covalentemente al hemo en el centro de cada subunidad. La unión de la primera molécula de oxígeno provoca un cambio conformacional en la hemoglobina que permite que la segunda molécula de oxígeno se una más fácilmente. A medida que cada molécula de oxígeno está unida, facilita aún más la unión de la siguiente molécula, hasta que los cuatro sitios hemo están ocupados por oxígeno. También ocurre lo contrario: Después de que la primera molécula de oxígeno se disocia y se “deja caer” en los tejidos, la siguiente molécula de oxígeno se disocia más fácilmente. Cuando los cuatro sitios hemo están ocupados, se dice que la hemoglobina está saturada. Cuando se ocupan de uno a tres sitios hemo, se dice que la hemoglobina está parcialmente saturada. Por lo tanto, al considerar la sangre como un todo, el porcentaje de las unidades hemo disponibles que están unidas al oxígeno en un momento dado se denomina saturación de hemoglobina. La saturación de hemoglobina del 100 por ciento significa que cada unidad hemo en todos los eritrocitos del cuerpo está unida al oxígeno. En un individuo sano con niveles normales de hemoglobina, la saturación de hemoglobina generalmente oscila entre 95 por ciento y 99 por ciento.
Disociación de oxígeno de la hemoglobina
La presión parcial es un aspecto importante de la unión del oxígeno y la disociación del hemo. Una curva de disociación oxígeno-hemoglobina es una gráfica que describe la relación de la presión parcial con la unión del oxígeno al hemo y su posterior disociación del hemo (Figura 3.2). Recuerde que los gases viajan de un área de mayor presión parcial a un área de menor presión parcial. Además, la afinidad de una molécula de oxígeno por el hemo aumenta a medida que se unen más moléculas de oxígeno. Por lo tanto, en la curva de saturación de oxígeno-hemoglobina, a medida que aumenta la presión parcial del oxígeno, un número proporcionalmente mayor de moléculas de oxígeno están unidas por el hemo. No es sorprendente que la curva de saturación/disociación de oxígeno-hemoglobina también muestre que cuanto menor sea la presión parcial de oxígeno, menos moléculas de oxígeno se unen al hemo. Como resultado, la presión parcial del oxígeno juega un papel importante en la determinación del grado de unión del oxígeno al hemo en el sitio de la membrana respiratoria, así como el grado de disociación del oxígeno del hemo en el sitio de los tejidos corporales.
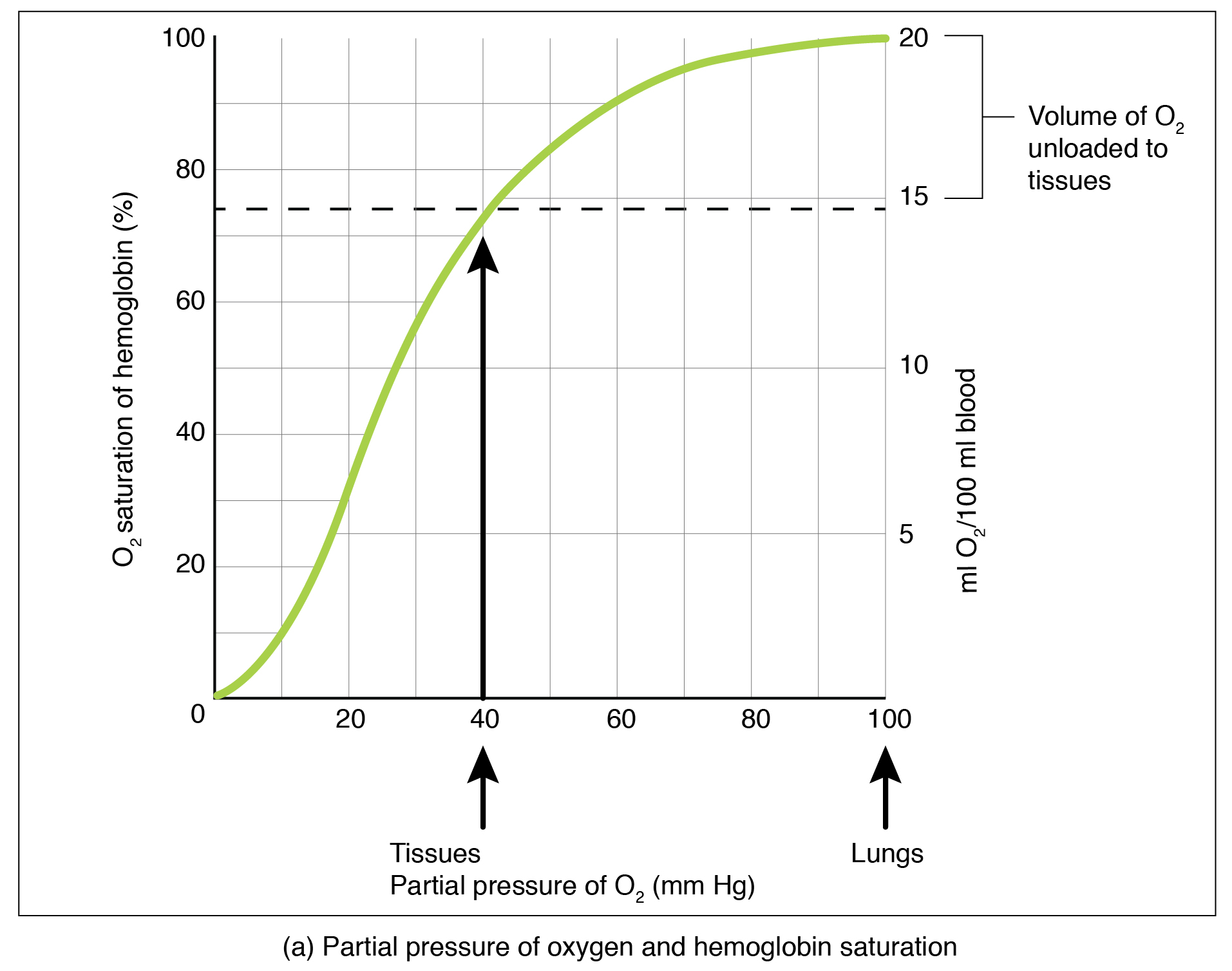


Figura 3.2. Disociación oxígeno-hemoglobina y efectos del pH y la temperatura. Estas tres gráficas muestran (a) la relación entre la presión parcial de oxígeno y la saturación de hemoglobina, (b) el efecto del pH sobre la curva de disociación oxígeno-hemoglobina, y (c) el efecto de la temperatura sobre la curva de disociación oxígeno-hemoglobina.
Los mecanismos detrás de la curva de saturación/disociación de oxígeno-hemoglobina también sirven como mecanismos de control automático que regulan la cantidad de oxígeno que se suministra a diferentes tejidos en todo el cuerpo. Esto es importante porque algunos tejidos tienen una tasa metabólica mayor que otros. Los tejidos altamente activos, como el músculo, utilizan rápidamente oxígeno para producir ATP, disminuyendo la presión parcial de oxígeno en el tejido a aproximadamente 20 mm Hg. La presión parcial del oxígeno dentro de los capilares es de aproximadamente 100 mm Hg, por lo que la diferencia entre los dos se vuelve bastante alta, alrededor de 80 mm Hg. Como resultado, un mayor número de moléculas de oxígeno se disocian de la hemoglobina y entran en los tejidos. Lo contrario es cierto para los tejidos, como el adiposo (grasa corporal), que tienen tasas metabólicas más bajas. Debido a que estas células utilizan menos oxígeno, la presión parcial de oxígeno dentro de dichos tejidos permanece relativamente alta, lo que resulta en menos moléculas de oxígeno que se disocian de la hemoglobina y entran en el fluido intersticial del tejido. Si bien se dice que la sangre venosa está desoxigenada, algo de oxígeno sigue unido a la hemoglobina en sus glóbulos rojos. Esto proporciona una reserva de oxígeno que se puede utilizar cuando los tejidos de repente demandan más oxígeno.
Factores distintos a la presión parcial también afectan la curva de saturación/disociación oxígeno-hemoglobina. Por ejemplo, una temperatura más alta promueve que la hemoglobina y el oxígeno se disocien más rápido, mientras que una temperatura más baja inhibe la disociación (ver Figura 3.2b). Sin embargo, el cuerpo humano regula fuertemente la temperatura, por lo que este factor puede no afectar el intercambio de gases en todo el cuerpo. La excepción a esto está en los tejidos altamente activos, que pueden liberar una mayor cantidad de energía de la que se emite como calor. Como resultado, el oxígeno se disocia fácilmente de la hemoglobina, que es un mecanismo que ayuda a proporcionar más oxígeno a los tejidos activos.
Ciertas hormonas, como los andrógenos, la epinefrina, las hormonas tiroideas y la hormona del crecimiento, pueden afectar la curva de saturación/disociación oxígeno-hemoglobina al estimular la producción de un compuesto llamado 2,3-bisfosfoglicerato (BPG) por los eritrocitos. El BPG es un subproducto de la glucólisis. Debido a que los eritrocitos no contienen mitocondrias, la glucólisis es el único método por el cual estas células producen ATP. El BPG promueve la disociación del oxígeno de la hemoglobina. Por lo tanto, cuanto mayor es la concentración de BPG, más fácilmente el oxígeno se disocia de la hemoglobina, a pesar de su presión parcial.
El pH de la sangre es otro factor que influye en la curva de saturación/disociación oxígeno-hemoglobina (ver Figura 3.2b). El efecto Bohr es un fenómeno que surge de la relación entre el pH y la afinidad del oxígeno por la hemoglobina: Un pH más bajo y más ácido promueve la disociación del oxígeno de la hemoglobina. En contraste, un pH más alto, o más básico, inhibe la disociación de oxígeno de la hemoglobina. Cuanto mayor sea la cantidad de dióxido de carbono en la sangre, más moléculas se deben convertir, lo que a su vez genera iones hidrógeno y así disminuye el pH de la sangre. Además, el pH de la sangre puede volverse más ácido cuando ciertos subproductos del metabolismo celular, como el ácido láctico, el ácido carbónico y el dióxido de carbono, se liberan en el torrente sanguíneo.
Transporte de dióxido de carbono en la sangre
El dióxido de carbono es transportado por tres mecanismos principales. El primer mecanismo de transporte de dióxido de carbono es por plasma sanguíneo, ya que algunas moléculas de dióxido de carbono se disuelven en la sangre. El segundo mecanismo es el transporte en forma de bicarbonato (HCO 3 —), que también se disuelve en plasma. El tercer mecanismo de transporte de dióxido de carbono es similar al transporte de oxígeno por los eritrocitos (Figura 3.3).

Figura 3.3. Transporte de dióxido de carbono. El dióxido de carbono es transportado por tres métodos diferentes: (a) en eritrocitos; (b) después de formar ácido carbónico (H2CO3), que se disuelve en plasma; (c) y en plasma.
Dióxido de carbono disuelto
Aunque no se considera que el dióxido de carbono sea altamente soluble en sangre, una pequeña fracción, aproximadamente del 7 al 10 por ciento, del dióxido de carbono que se difunde a la sangre desde los tejidos se disuelve en plasma. El dióxido de carbono disuelto luego viaja en el torrente sanguíneo y cuando la sangre llega a los capilares pulmonares, el dióxido de carbono disuelto se difunde a través de la membrana respiratoria hacia los alvéolos, donde luego se exhala durante la ventilación pulmonar.
Buffer de bicarbonato
Una gran fracción, alrededor del 70 por ciento, de las moléculas de dióxido de carbono que se difunden en la sangre se transporta a los pulmones como bicarbonato. La mayor parte del bicarbonato se produce en los eritrocitos después de que el dióxido de carbono se difunde en los capilares y posteriormente en los glóbulos rojos. La anhidrasa carbónica (CA) provoca que el dióxido de carbono y el agua formen ácido carbónico (H 2 CO 3), que se disocia en dos iones: bicarbonato (HCO 3 —) e hidrógeno (H +). La siguiente fórmula representa esta reacción:
\[\ce{CO2 + H2O CA↔ H2CO3↔H+ + HCO3−CO2 + H2O CA↔ H2CO3↔H+ + HCO3−}\]
El bicarbonato tiende a acumularse en los eritrocitos de manera que hay una mayor concentración de bicarbonato en los eritrocitos que en el plasma sanguíneo circundante. Como resultado, parte del bicarbonato saldrá de los eritrocitos y bajará su gradiente de concentración hacia el plasma a cambio de iones cloruro (Cl —). A este fenómeno se le conoce como el desplazamiento de cloruro y se produce porque al intercambiar un ion negativo por otro ion negativo, no se altera ni la carga eléctrica de los eritrocitos ni la de la sangre.
En los capilares pulmonares, se invierte la reacción química que produjo el bicarbonato (mostrada arriba), y el dióxido de carbono y el agua son los productos. Gran parte del bicarbonato en el plasma vuelve a entrar en los eritrocitos a cambio de iones cloruro. Los iones hidrógeno y los iones bicarbonato se unen para formar ácido carbónico, que es convertido en dióxido de carbono y agua por la anhidrasa carbónica. El dióxido de carbono se difunde fuera de los eritrocitos y hacia el plasma, donde puede difunderse aún más a través de la membrana respiratoria hacia los alvéolos para ser exhalado durante la ventilación pulmonar.
Carbaminohemoglobina
Alrededor del 20 por ciento del dióxido de carbono está unido por la hemoglobina y es transportado a los pulmones. El dióxido de carbono no se une al hierro como lo hace el oxígeno; en cambio, el dióxido de carbono une restos de aminoácidos en las porciones de globina de la hemoglobina para formar carbaminohemoglobina, que se forma cuando la hemoglobina y el dióxido de carbono se unen. Cuando la hemoglobina no está transportando oxígeno, tiende a tener un tono azulado púrpura, creando el color granate más oscuro típico de la sangre desoxigenada. La siguiente fórmula representa esta reacción reversible:
\[\ce{CO2 + Hb↔HbCO2CO2 + Hb↔HbCO2}\]
Similar al transporte de oxígeno por hemo, la unión y disociación del dióxido de carbono hacia y desde la hemoglobina depende de la presión parcial del dióxido de carbono. Debido a que el dióxido de carbono se libera de los pulmones, la sangre que sale de los pulmones y llega a los tejidos del cuerpo tiene una presión parcial de dióxido de carbono menor que la que se encuentra en los tejidos. Como resultado, el dióxido de carbono sale de los tejidos debido a su mayor presión parcial, ingresa a la sangre, y luego se mueve hacia los glóbulos rojos, uniéndose a la hemoglobina. En contraste, en los capilares pulmonares, la presión parcial del dióxido de carbono es alta en comparación con dentro de los alvéolos. Como resultado, el dióxido de carbono se disocia fácilmente de la hemoglobina y se difunde a través de la membrana respiratoria hacia el aire.
Además de la presión parcial del dióxido de carbono, la saturación de oxígeno de la hemoglobina y la presión parcial de oxígeno en la sangre también influyen en la afinidad de la hemoglobina por el dióxido de carbono. El efecto Haldane es un fenómeno que surge de la relación entre la presión parcial del oxígeno y la afinidad de la hemoglobina por el dióxido de carbono. La hemoglobina que está saturada de oxígeno no se une fácilmente al dióxido de carbono. Sin embargo, cuando el oxígeno no está unido al hemo y la presión parcial del oxígeno es baja, la hemoglobina se une fácilmente al dióxido de carbono.
notas
- Observe esto para ver el transporte de oxígeno de los pulmones a los tejidos.
Ejercicio\(\PageIndex{1}\)
La oxihemoglobina se forma por una reacción química entre ¿cuál de las siguientes?
a. hemoglobina y dióxido de carbono
b. anhidrasa carbónica y dióxido de carbono
c. hemoglobina y oxígeno
d. anhidrasa carbónica y oxígeno
Ejercicio\(\PageIndex{2}\)
¿Cuál de los siguientes factores juega un papel en la curva de saturación/disociación oxígeno-hemoglobina?
a. temperatura
b. pH
c. BPG
d. Todo lo anterior
Ejercicio\(\PageIndex{3}\)
¿Cuál de las siguientes situaciones ocurre durante el cambio de cloruro?
a. El cloruro se elimina del eritrocito.
b. El cloruro se cambia por bicarbonato.
c. El bicarbonato se elimina del eritrocito.
d. El bicarbonato se elimina de la sangre
Ejercicio\(\PageIndex{4}\)
Una baja presión parcial de oxígeno promueve la unión de la hemoglobina al dióxido de carbono. Este es un ejemplo del ________.
a. Efecto Haldane
b. Efecto Bohr
c. Ley de Dalton
d. Ley de Henry
Ejercicio\(\PageIndex{5}\)
¿Por qué la sangre oxigenada es de color rojo brillante, mientras que la sangre desoxigenada tiende a ser más de color púrpura? (piensa en el video sobre el transporte de oxígeno)
Ejercicio\(\PageIndex{6}\)
Describir la relación entre la presión parcial del oxígeno y la unión del oxígeno a la hemoglobina.