
9.2: Vías de señalización, hormonas y sistema endocrino
- Page ID
- 121075
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Objetivos de aprendizaje
- Describir estrategias generales de señalización celular y explicar los tipos de hormonas y sus vías de señalización en la comunicación endocrina
Señalización química
Hay dos tipos de comunicación en el mundo de las células vivas. La comunicación entre células se llama señalización intercelular, y la comunicación dentro de una célula se llama señalización intracelular. Una manera fácil de recordar la distinción es entendiendo el origen latino de los prefijos: inter- significa “entre” (por ejemplo, las líneas que se cruzan son las que se cruzan entre sí) e intra- significa “dentro” (como intravenosa).
Las señales químicas son liberadas por las células de señalización en forma de moléculas pequeñas, generalmente volátiles o solubles llamadas ligandos. Un ligando es una molécula que se une a otra molécula específica, en algunos casos, entregando una señal en el proceso. Por lo tanto, los ligandos pueden considerarse como moléculas de señalización. Los ligandos interactúan con las proteínas en las células diana, que son células que se ven afectadas por señales químicas; estas proteínas también se denominan receptores. Los ligandos y receptores existen en varias variedades; sin embargo, un ligando específico tendrá un receptor específico que típicamente se une solo a ese ligando.
Formas de Señalización
Hay cuatro categorías de señalización química que se encuentran en organismos multicelulares: señalización paracrina, señalización endocrina, señalización autocrina y señalización directa a través de uniones gap (Figura 9.2). La principal diferencia entre las diferentes categorías de señalización es la distancia que recorre la señal a través del organismo para llegar a la célula diana. No todas las células se ven afectadas por las mismas señales.

Figura 9.2. Formas de señalización química: autocrinas, uniones gap, paracrinas y endocrinas.
En la señalización química, una célula puede dirigirse a sí misma (señalización autocrina), una célula conectada por uniones de brecha, una célula cercana (señalización paracrina) o una célula distante (señalización endocrina). La señalización paracrina actúa sobre las células cercanas, la señalización endocrina usa el sistema circulatorio para transportar ligandos y la señalización autocrina actúa sobre la célula de señalización. La señalización a través de uniones gap implica moléculas de señalización que se mueven directamente entre células adyacentes.
Señalización paracrina
Las señales que actúan localmente entre células que están muy juntas se denominan señales paracrinas. Las señales paracrinas se mueven por difusión a través de la matriz extracelular. Este tipo de señales suelen provocar respuestas rápidas que duran solo un corto período de tiempo. Para mantener la respuesta localizada, las moléculas de ligando paracrino normalmente se degradan rápidamente por las enzimas o se eliminan por las células vecinas. Al eliminar las señales se restablecerá el gradiente de concentración de la señal, permitiendo que se difundan rápidamente a través del espacio intracelular si se liberan de nuevo.
Un ejemplo de señalización paracrina es la transferencia de señales a través de sinapsis entre células nerviosas. Una célula nerviosa consiste en un cuerpo celular, varias extensiones cortas y ramificadas llamadas dendritas que reciben estímulos, y una extensión larga llamada axón, que transmite señales a otras células nerviosas o células musculares. La unión entre las células nerviosas donde ocurre la transmisión de señales se llama sinapsis. Una señal sináptica es una señal química que viaja entre las células nerviosas. Las señales dentro de las células nerviosas se propagan por impulsos eléctricos de rápido movimiento. Cuando estos impulsos alcanzan el final del axón, la señal continúa a una dendrita de la siguiente célula por la liberación de ligandos químicos llamados neurotransmisores por la célula presináptica (la célula que emite la señal). Los neurotransmisores son transportados a través de las distancias muy pequeñas entre las células nerviosas, las cuales se denominan sinapsis químicas (Figura 9.3). La pequeña distancia entre las células nerviosas permite que la señal viaje rápidamente; esto permite una respuesta inmediata, como, ¡Quita la mano de la estufa!
Cuando el neurotransmisor se une al receptor en la superficie de la célula postsináptica, el potencial electroquímico de la célula diana cambia y se lanza el siguiente impulso eléctrico. Los neurotransmisores que se liberan en la sinapsis química se degradan rápidamente o son reabsorbidos por la célula presináptica para que la célula nerviosa receptora pueda recuperarse rápidamente y estar preparada para responder rápidamente a la siguiente señal sináptica.

Figura 9.3. Sinapsis que muestra liberación de neurotransmisores.
La distancia entre la célula presináptica y la célula postsináptica, llamada brecha sináptica, es muy pequeña y permite una rápida difusión del neurotransmisor. Las enzimas en la hendidura sinapática degradan algunos tipos de neurotransmisores para terminar la señal.
Señalización endocrina
Las señales de células distantes se llaman señales endocrinas y se originan a partir de células endocrinas. (En el cuerpo, muchas células endocrinas se encuentran en las glándulas endocrinas, como la glándula tiroides, el hipotálamo y la glándula pituitaria). Este tipo de señales suelen producir una respuesta más lenta pero tienen un efecto más duradero. Los ligandos liberados en la señalización endocrina se denominan hormonas, moléculas de señalización que se producen en una parte del cuerpo pero que afectan a otras regiones del cuerpo a cierta distancia.
Las hormonas recorren las grandes distancias entre las células endocrinas y sus células diana a través del torrente sanguíneo, que es una forma relativamente lenta de moverse por todo el cuerpo. Por su forma de transporte, las hormonas se diluyen y están presentes en bajas concentraciones cuando actúan sobre sus células diana. Esto es diferente de la señalización paracrina, en la que las concentraciones locales de ligandos pueden ser muy altas.
Señalización autocrina
Las señales autocrinas son producidas por células de señalización que también pueden unirse al ligando que se libera. Esto significa que la célula de señalización y la célula de destino pueden ser la misma o una célula similar (el prefijo auto- significa auto, un recordatorio de que la célula de señalización envía una señal a sí misma). Este tipo de señalización suele ocurrir durante el desarrollo temprano de un organismo para asegurar que las células se desarrollen en los tejidos correctos y asuman la función adecuada. La señalización autocrina también regula la sensación de dolor y las respuestas inflamatorias. Además, si una célula está infectada con un virus, la célula puede señalarse a sí misma para que sufra una muerte celular programada, matando al virus en el proceso. En algunos casos, las células vecinas del mismo tipo también están influenciadas por el ligando liberado. En el desarrollo embriológico, este proceso de estimulación de un grupo de células vecinas puede ayudar a dirigir la diferenciación de células idénticas en el mismo tipo celular, asegurando así el resultado adecuado del desarrollo.
Señalización directa a través de uniones de brecha
Las uniones de brecha en animales y plasmodesmas en plantas son conexiones entre las membranas plasmáticas de las células vecinas. Estos canales llenos de agua permiten que pequeñas moléculas de señalización, llamadas mediadores intracelulares, se difundan entre las dos células. Las moléculas pequeñas, como los iones calcio (Ca2+), son capaces de moverse entre las células, pero las moléculas grandes como las proteínas y el ADN no pueden caber a través de los canales. La especificidad de los canales asegura que las células permanezcan independientes pero puedan transmitir señales rápida y fácilmente. La transferencia de moléculas de señalización comunica el estado actual de la célula que está directamente al lado de la célula diana; esto permite que un grupo de células coordine su respuesta a una señal que solo una de ellas pudo haber recibido. En las plantas, los plasmodesmas son ubicuos, convirtiendo a toda la planta en una red de comunicación gigante.
Tipos de Receptores
Los receptores son moléculas proteicas en la célula diana o en su superficie que se unen al ligando. Existen dos tipos de receptores, receptores internos y receptores de superficie celular.
Receptores internos
Los receptores internos, también conocidos como receptores intracelulares o citoplásmicos, se encuentran en el citoplasma de la célula y responden a moléculas de ligando hidrófobas que son capaces de viajar a través de la membrana plasmática. Una vez dentro de la célula, muchas de estas moléculas se unen a proteínas que actúan como reguladores de la síntesis de ARNm (transcripción) para mediar en la expresión génica. La expresión génica es el proceso celular de transformar la información en el ADN de una célula en una secuencia de aminoácidos, que finalmente forma una proteína. Cuando el ligando se une al receptor interno, se desencadena un cambio conformacional que expone un sitio de unión a ADN en la proteína. El complejo ligando-receptor se mueve hacia el núcleo, luego se une a regiones reguladoras específicas del ADN cromosómico y promueve el inicio de la transcripción (Figura 9.4). La transcripción es el proceso de copiar la información en el ADN de una célula en una forma especial de ARN llamada ARN mensajero (ARNm); la célula utiliza información en el ARNm (que se mueve hacia el citoplasma y se asocia con ribosomas) para enlazar aminoácidos específicos en el orden correcto, produciendo una proteína. Los receptores internos pueden influir directamente en la expresión génica sin tener que pasar la señal a otros receptores o mensajeros.

Las moléculas de señalización hidrofóbicas normalmente se difunden a través de la membrana plasmática e interactúan con los receptores intracelulares Muchos receptores intracelulares son factores de transcripción que interactúan con el ADN en el núcleo y regulan la expresión génica.
Receptores de superficie celular
Los receptores de superficie celular, también conocidos como receptores transmembrana, son proteínas de superficie celular ancladas a membrana (integrales) que se unen a moléculas de ligando externas. Este tipo de receptor abarca la membrana plasmática y realiza la transducción de señales, en la que una señal extracelular se convierte en una señal intercelular. Los ligandos que interactúan con los receptores de la superficie celular no tienen que ingresar a la célula a la que afectan. Los receptores de superficie celular también se denominan proteínas o marcadores específicos de células porque son específicos de tipos celulares individuales.
Debido a que las proteínas receptoras de la superficie celular son fundamentales para el funcionamiento normal de las células, no debería sorprender que un mal funcionamiento en alguna de estas proteínas pueda tener graves consecuencias. Se ha demostrado que los errores en las estructuras proteicas de ciertas moléculas receptoras juegan un papel en la hipertensión (presión arterial alta), el asma, las enfermedades cardíacas y el cáncer.
Cada receptor de superficie celular tiene tres componentes principales: un dominio externo de unión a ligando, una región hidrofóbica que abarca la membrana y un dominio intracelular dentro de la célula. El dominio de unión a ligando también se denomina dominio extracelular. El tamaño y extensión de cada uno de estos dominios varían ampliamente, dependiendo del tipo de receptor. Los receptores de superficie celular están involucrados en la mayor parte de la señalización en organismos multicelulares. Hay tres categorías generales de receptores de superficie celular: receptores unidos a canales iónicos, receptores unidos a proteínas G y receptores ligados a enzimas.
Los receptores unidos a canales iónicos se unen a un ligando y abren un canal a través de la membrana que permite el paso de iones específicos. Para formar un canal, este tipo de receptor de superficie celular tiene una extensa región que abarca la membrana. Para interactuar con las colas de ácidos grasos fosfolípidos que forman el centro de la membrana plasmática, muchos de los aminoácidos en la región que abarca la membrana son de naturaleza hidrófoba. Por el contrario, los aminoácidos que recubren el interior del canal son hidrófilos para permitir el paso de agua o iones. Cuando un ligando se une a la región extracelular del canal, hay un cambio conformacional en la estructura de las proteínas que permite el paso de iones como sodio, calcio, magnesio e hidrógeno (Figura 9.5)

Figura 9.5. Un canal iónico cerrado. Los canales iónicos cerrados forman un poro a través de la membrana plasmática que se abre cuando la molécula de señalización se une. El poro abierto permite entonces que los iones fluyan dentro o fuera de la celda.
Los receptores unidos a proteínas G se unen a un ligando y activan una proteína de membrana llamada proteína G. La proteína G activada interactúa entonces con un canal iónico o una enzima en la membrana (Figura 9.6). Todos los receptores unidos a la proteína G tienen siete dominios transmembrana, pero cada receptor tiene su propio dominio extracelular específico y el sitio de unión a la proteína G.
La señalización celular usando receptores unidos a proteína G ocurre como una serie cíclica de eventos. Antes de que el ligando se una, la proteína G inactiva puede unirse a un sitio recientemente revelado en el receptor específico para su unión. Una vez que la proteína G se une al receptor, el cambio de forma resultante activa la proteína G, que libera GDP y recoge GTP. Las subunidades de la proteína G se dividieron en la subunidad α y la subunidad βγ. Uno o ambos de estos fragmentos de proteína G pueden ser capaces de activar otras proteínas como resultado. Después de un tiempo, el GTP en la subunidad α activa de la proteína G se hidroliza al PIB y la subunidad βγ se desactiva. Las subunidades se reasocian para formar la proteína G inactiva y el ciclo comienza de nuevo.
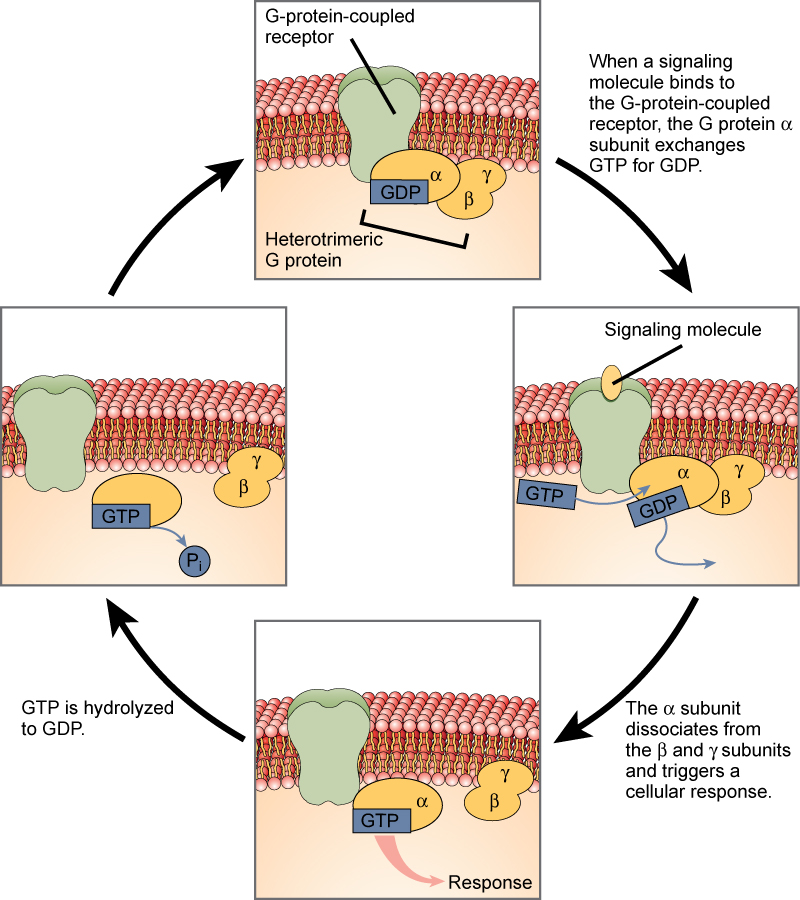
Figura 9.6. Las proteínas G heterotriméricas tienen tres subunidades: α, β y γ. Cuando una molécula de señalización se une a un receptor acoplado a proteína G en la membrana plasmática, una molécula GDP asociada con la subunidad α se intercambia por GTP. Las subunidades β y γ se disocian de la subunidad α, y una respuesta celular es desencadenada por la subunidad α o por el par βγ disociado. La hidrólisis de GTP a PIB termina la señal.
Los receptores ligados a proteínas G han sido ampliamente estudiados y se ha aprendido mucho sobre su papel en el mantenimiento de la salud. Las bacterias que son patógenas para los humanos pueden liberar venenos que interrumpen la función específica del receptor ligado a la proteína G, lo que lleva a enfermedades como la tos ferina, el botulismo y el cólera. En el cólera (Figura 9.7), por ejemplo, la bacteria transportada por el agua Vibrio cholerae produce una toxina, coleragen, que se une a las células que recubren el intestino delgado. La toxina luego ingresa a estas células intestinales, donde modifica una proteína G que controla la apertura de un canal de cloruro y hace que permanezca continuamente activa, resultando en grandes pérdidas de líquidos del organismo y como consecuencia de ello una deshidratación potencialmente fatal.

Figura 9.7. Transmitido principalmente a través del agua potable contaminada, el cólera es una de las principales causas de muerte en el mundo en desarrollo y en áreas donde los desastres naturales interrumpen la disponibilidad de agua potable La bacteria del cólera, Vibrio cholerae, crea una toxina que modifica las vías de señalización celular mediadas por proteína G en los intestinos. El saneamiento moderno elimina la amenaza de brotes de cólera, como el que arrasó la ciudad de Nueva York en 1866. Este cartel de esa época muestra cómo, en ese momento, no se entendía la forma en que se transmitió la enfermedad. (crédito: Comisión Sanitaria de la Ciudad de Nueva York)
Los receptores ligados a enzimas son receptores de superficie celular con dominios intracelulares que están asociados con una enzima. En algunos casos, el dominio intracelular del receptor en sí es una enzima. Otros receptores ligados a enzimas tienen un pequeño dominio intracelular que interactúa directamente con una enzima. Los receptores ligados a enzimas normalmente tienen grandes dominios extracelulares e intracelulares, pero la región que abarca la membrana consiste en una única región alfa-helicoidal de la cadena peptídica. Cuando un ligando se une al dominio extracelular, se transfiere una señal a través de la membrana, activando la enzima. La activación de la enzima desencadena una cadena de eventos dentro de la célula que eventualmente conduce a una respuesta. Un ejemplo de este tipo de receptor ligado a enzimas es el receptor de tirosina quinasa (Figura 9.8). Una quinasa es una enzima que transfiere grupos fosfato de ATP a otra proteína. El receptor de tirosina quinasa transfiere grupos fosfato a moléculas de tirosina (residuos de tirosina). Primero, las moléculas de señalización se unen al dominio extracelular de dos receptores cercanos de tirosina quinasa. Los dos receptores vecinos se unen o se dimerizan. Luego se agregan fosfatos a los residuos de tirosina en el dominio intracelular de los receptores (fosforilación). Los residuos fosforilados pueden entonces transmitir la señal al siguiente mensajero dentro del citoplasma.

Figura 9.8. Un receptor tirosina quinasa es un receptor ligado a enzimas con una única región transmembrana y dominios extracelulares e intracelulares. La unión de una molécula de señalización al dominio extracelular hace que el receptor se dimerice. Los residuos de tirosina en el dominio intracelular se autofosforilan, desencadenando una respuesta celular aguas abajo. La señal es terminada por una fosfatasa que elimina los fosfatos de los residuos de fosfotirosina.
Moléculas de señalización
Producidos por las células de señalización y la posterior unión a los receptores en las células diana, los ligandos actúan como señales químicas que viajan a las células diana para coordinar las respuestas. Los tipos de moléculas que sirven como ligandos son increíblemente variados y van desde pequeñas proteínas hasta pequeños iones como el calcio (Ca2+).
Ligandos hidrófobos pequeños
Los ligandos hidrófobos pequeños pueden difundirse directamente a través de la membrana plasmática e interactuar con receptores internos. Miembros importantes de esta clase de ligandos son las hormonas esteroides. Los esteroides son lípidos que tienen un esqueleto hidrocarbonado con cuatro anillos fusionados; diferentes esteroides tienen diferentes grupos funcionales unidos al esqueleto de carbono. Las hormonas esteroides incluyen la hormona sexual femenina, estradiol, que es un tipo de estrógeno; la hormona sexual masculina, la testosterona; y el colesterol, que es un componente estructural importante de las membranas biológicas y un precursor de las hormonas estéridas (Figura 9.9). Otras hormonas hidrofóbicas incluyen las hormonas tiroideas y la vitamina D. Para ser solubles en sangre, los ligandos hidrófobos deben unirse a proteínas portadoras mientras se transportan a través del torrente sanguíneo.
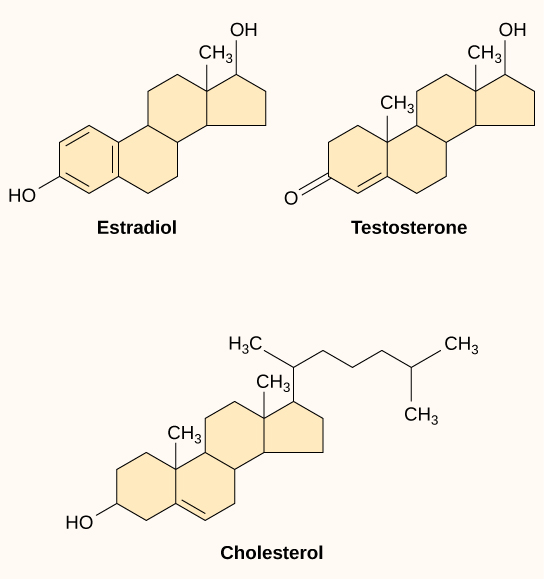
Figura 9.9. Las hormonas esteroides tienen estructuras químicas similares a su precursor, el colesterol. Debido a que estas moléculas son pequeñas e hidrofóbicas, pueden difundirse directamente a través de la membrana plasmática hacia la célula, donde interactúan con los receptores internos.
Ligandos solubles en agua
Los ligandos solubles en agua son polares y por lo tanto no pueden pasar a través de la membrana plasmática sin ayuda; a veces, son demasiado grandes para pasar a través de la membrana en absoluto. En cambio, la mayoría de los ligandos solubles en agua se unen al dominio extracelular de los receptores de superficie celular Este grupo de ligandos es bastante diverso e incluye moléculas pequeñas, péptidos y proteínas.
Otros ligandos
El óxido nítrico (NO) es un gas que también actúa como ligando. Es capaz de difundirse directamente a través de la membrana plasmática, y una de sus funciones es interactuar con los receptores en el músculo liso e inducir la relajación del tejido. El NO tiene una vida media muy corta y por lo tanto solo funciona a distancias cortas. La nitroglicerina, un tratamiento para enfermedades cardíacas, actúa desencadenando la liberación de NO, lo que hace que los vasos sanguíneos se dilaten (expandan), restaurando así el flujo sanguíneo al corazón. El NO se ha vuelto más conocido recientemente porque la vía que afecta está dirigida por medicamentos recetados para la disfunción eréctil, como el Viagra (la erección implica vasos sanguíneos dilatados).
Hormonas
Las hormonas median los cambios en las células diana al unirse a receptores hormonales específicos. De esta manera, a pesar de que las hormonas circulan por todo el cuerpo y entran en contacto con muchos tipos celulares diferentes, solo afectan a las células que poseen los receptores necesarios. Los receptores para una hormona específica pueden encontrarse en muchas células diferentes o pueden limitarse a un pequeño número de células especializadas. Por ejemplo, las hormonas tiroideas actúan sobre muchos tipos de tejidos diferentes, estimulando la actividad metabólica en todo el cuerpo. Las células pueden tener muchos receptores para la misma hormona pero a menudo también poseen receptores para diferentes tipos de hormonas. El número de receptores que responden a una hormona determina la sensibilidad de la célula a esa hormona, y la respuesta celular resultante. Adicionalmente, el número de receptores que responden a una hormona puede cambiar con el tiempo, resultando en aumento o disminución de la sensibilidad celular. En la regulación positiva, el número de receptores aumenta en respuesta al aumento de los niveles hormonales, haciendo que la célula sea más sensible a la hormona y permitiendo una mayor actividad celular. Cuando el número de receptores disminuye en respuesta al aumento de los niveles hormonales, llamados regulación a la baja, la actividad celular se reduce.
La unión al receptor altera la actividad celular y da como resultado un aumento o disminución de los procesos corporales normales. Dependiendo de la ubicación del receptor de proteínas en la célula diana y la estructura química de la hormona, las hormonas pueden mediar los cambios directamente al unirse a los receptores hormonales intracelulares y modular la transcripción génica, o indirectamente al unirse a los receptores de la superficie celular y estimular vías de señalización.
Receptores hormonales intracelulares
Las hormonas derivadas de lípidos (solubles), como las hormonas esteroides, se difunden a través de las membranas de la célula endocrina. Una vez fuera de la célula, se unen para transportar proteínas que las mantienen solubles en el torrente sanguíneo. En la célula diana, las hormonas se liberan de la proteína portadora y se difunden a través de la bicapa lipídica de la membrana plasmática de las células. Las hormonas esteroides pasan a través de la membrana plasmática de una célula diana y se adhieren a receptores intracelulares que residen en el citoplasma o en el núcleo. Las vías de señalización celular inducidas por las hormonas esteroides regulan genes específicos en el ADN de la célula. Las hormonas y el complejo receptor actúan como reguladores de la transcripción al aumentar o disminuir la síntesis de moléculas de ARNm de genes específicos. Esto, a su vez, determina la cantidad de proteína correspondiente que se sintetiza alterando la expresión génica. Esta proteína puede ser utilizada ya sea para cambiar la estructura de la célula o para producir enzimas que catalizan reacciones químicas. De esta manera, la hormona esteroidea regula procesos celulares específicos como se ilustra en la Figura 9.1.
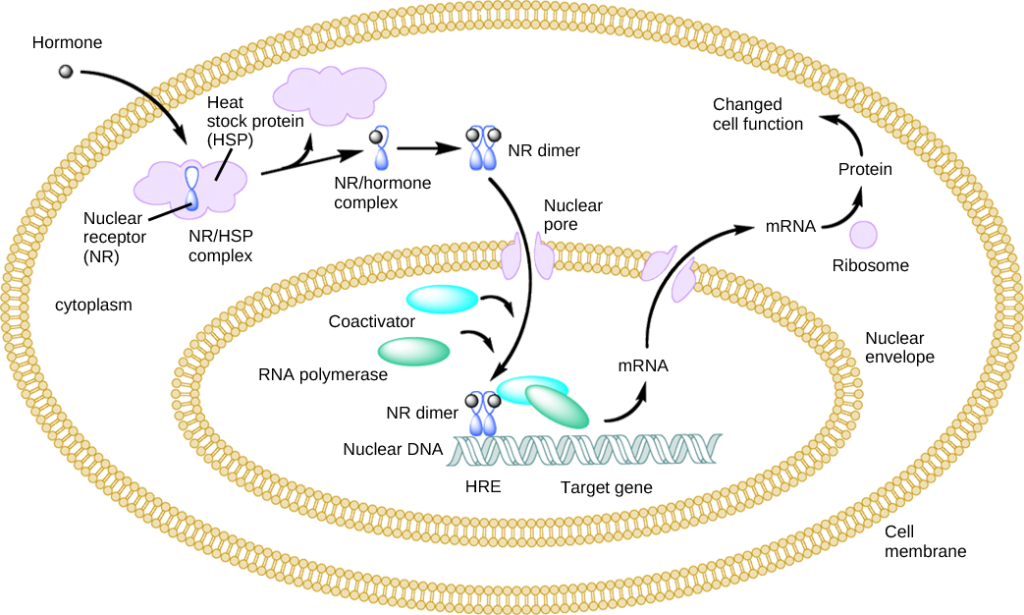
Otras hormonas liposolubles que no son hormonas esteroides, como la vitamina D y la tiroxina, tienen receptores localizados en el núcleo. Las hormonas se difunden tanto a través de la membrana plasmática como de la envoltura nuclear, luego se unen a los receptores en el núcleo. El complejo hormona-receptor estimula la transcripción de genes específicos.
Receptores hormonales de membrana plasmática
Las hormonas derivadas de aminoácidos y las hormonas polipeptídicas no son derivadas de lípidos (solubles en lípidos) y por tanto no pueden difundirse a través de la membrana plasmática de las células. Las hormonas insolubles en lípidos se unen a receptores en la superficie externa de la membrana plasmática, a través de receptores hormonales de membrana plasmática. A diferencia de las hormonas esteroides, las hormonas insolubles en lípidos no afectan directamente a la célula diana porque no pueden ingresar a la célula y actuar directamente sobre el ADN. La unión de estas hormonas a un receptor de superficie celular resulta en la activación de una vía de señalización; esto desencadena la actividad intracelular y lleva a cabo los efectos específicos asociados a la hormona. De esta manera, nada pasa a través de la membrana celular; la hormona que se une en la superficie permanece en la superficie de la célula mientras que el producto intracelular permanece dentro de la célula. La hormona que inicia la vía de señalización se denomina primer mensajero, que activa un segundo mensajero en el citoplasma, como se ilustra en la Figura 9.6.
Un segundo mensajero muy importante es el AMP cíclico (AMPc). Cuando una hormona se une a su receptor de membrana, se activa una proteína G que se asocia con el receptor; las proteínas G son proteínas separadas de los receptores que se encuentran en la membrana celular. Cuando una hormona no está unida al receptor, la proteína G es inactiva y se une al difosfato de guanosina, o GDP. Cuando una hormona se une al receptor, la proteína G se activa al unirse al trifosfato de guanosina, o GTP, en lugar del PIB. Después de la unión, el GTP es hidrolizado por la proteína G en GDP y se vuelve inactiva.La proteína G activada a su vez activa una enzima unida a la membrana llamada adenilil ciclasa. La adenilciclasa cataliza la conversión de ATP en AMPc. El cAMP, a su vez, activa un grupo de proteínas llamadas proteínas quinasas, que transfieren un grupo fosfato de ATP a una molécula sustrato en un proceso llamado fosforilación. La fosforilación de una molécula sustrato cambia su orientación estructural, activándola así. Estas moléculas activadas pueden entonces mediar cambios en los procesos celulares.El efecto de una hormona se amplifica a medida que avanza la vía de señalización. La unión de una hormona a un solo receptor provoca la activación de muchas proteínas G, lo que activa la adenil ciclasa. Cada molécula de adenil ciclasa desencadena entonces la formación de muchas moléculas de AMPc. La amplificación adicional ocurre ya que las proteínas quinasas, una vez activadas por AMPc, pueden catalizar muchas reacciones. De esta manera, una pequeña cantidad de hormona puede desencadenar la formación de una gran cantidad de producto celular. Para detener la actividad hormonal, el AMPc es desactivado por la enzima citoplasmática fosfodiesterasa, o PDE. La PDE siempre está presente en la célula y descompone el AMPc para controlar la actividad hormonal, evitando la sobreproducción de productos celulares.La respuesta específica de una célula a una hormona insoluble en lípidos depende del tipo de receptores que están presentes en la membrana celular y las moléculas de sustrato presentes en la célula citoplasma. Las respuestas celulares a la unión hormonal de un receptor incluyen alterar la permeabilidad de la membrana y las vías metabólicas, estimular la síntesis de proteínas y enzimas, y activar la liberación de hormonas.
Ejercicio\(\PageIndex{1}\)
Se ha descubierto una nueva molécula antagonista que se une y bloquea los receptores de membrana plasmática. ¿Qué efecto tendrá este antagonista sobre la testosterona, una hormona esteroide?
a. Bloqueará la unión de la testosterona a su receptor.
b. Bloqueará la testosterona para que no active la señalización de cAMP.
c. Aumentará la señalización mediada por testosterona.
d. No afectará la señalización mediada por testosterona.
Ejemplo\(\PageIndex{2}\):
¿Qué efecto tendrá un inhibidor de AMPc en una vía de señalización mediada por hormonas peptídicas?
a. Evitará que la hormona se una a su receptor.
b. Evitará la activación de una proteína G.
c. Evitará la activación de la adenilato ciclasa.
d. Evitará la activación de proteínas quinasas.
Ejercicio\(\PageIndex{3}\)
Nombrar dos funciones importantes de los receptores hormonales.
Ejercicio\(\PageIndex{4}\)
¿Cómo pueden las hormonas mediar los cambios?
Ejercicio\(\PageIndex{5}\)
Enumere todas las estructuras que intervienen en la señalización hormonal general. Una vez que tenga todas las estructuras listadas, organícelas de pequeñas a grandes. A continuación, organícelos en orden de eventos durante la señalización hormonal, de la primera a la última.