
1.1: Revisión de Biología Celular
- Page ID
- 121777

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
- Descripción general de la biología celular
- Dentro de la celda:
- Membrana celular, citoplasma
- Organelos
- Fuera de la celda:
- Matriz extra-celular (ECM)
- Fluido Extracelular (ECF)
- Ni dentro ni fuera:
- División celular
- Muerte celular
- Unión celular

Descripción general de la biología celular
La célula es la unidad básica de la vida. Es lo más pequeño que llamamos vivir (sin discutir sobre virus), y el cuerpo humano está hecho de 10 billones de ellos. Empezamos como una sola célula, y el propósito de esta clase fue aprender un poco sobre cómo esa célula se convirtió en el organismo trillón celular que eres hoy. Casi todas las instrucciones para hacer billones de células —cómo hacerlas, cuándo hacerlas, dónde hacerlas— se encuentran dentro de esa celda única. Diferentes células tienen diferentes funciones, aunque casi todas las células de una persona tienen exactamente el mismo ADN. Para diferenciarse unas de otras, las células expresan ADN diferente. Este es un proceso importante llamado diferenciación, que en realidad no es más complicado en concepto que las células que van de parecer células genéricas aburridas a verse y comportarse de manera diferente a otras células. Damos diferentes nombres a las celdas a medida que hacen estos cambios. Por ejemplo, una célula madre que da origen a, oh digamos, un pitufo, se llamaría Célula Madre Pitufal. A medida que las Células Madre Pitufales se dividen y se diferencian en células que hacen un pitufo, las células que realmente hacen el pitufo se llamarían Pitufoblastos, y cuando el Pitufo estuviera terminado, las células dentro de ese Pitufo se llamarían Pitufocitos. Las células madre que se encuentran en un tejido llamado mesénquima se denominan células madre mesenquimales, las células que producen la dentina se denominan odontoblastos, las células dentro del cemento maduro se denominan cementocitos. Nótese que la diferenciación no es un solo paso, sino una serie de pasos a lo largo de un espectro, desde los menos diferenciados, hasta los más diferenciados, hasta llegar a una célula diferenciada terminalmente. El objetivo de este capítulo es revisar aspectos de una clase de biología celular que aparecen en este libro de texto, y no mucho más. Asumimos que ha cubierto la biología celular en un por-requisito para esta clase. Necesitamos revisar las partes de una célula que nos ayuden a explicar conceptos difíciles como la diferenciación y el desarrollo. Si encuentras que necesitas algo más que un repaso rápido, aquí tienes algún enlace de los NIH, quienes cuentan con una serie de publicaciones y videos muy útiles (gratis), como:
- Animaciones 3D de la célula humana
- Aprender genética (sitio web interactivo)
- Dentro de la celda (descargas pdf y epub)
Dos libros electrónicos útiles (y gratuitos) que tal vez desee descargar son de OpenStax:

Dentro de la celda
Membrana celular y citoplasma
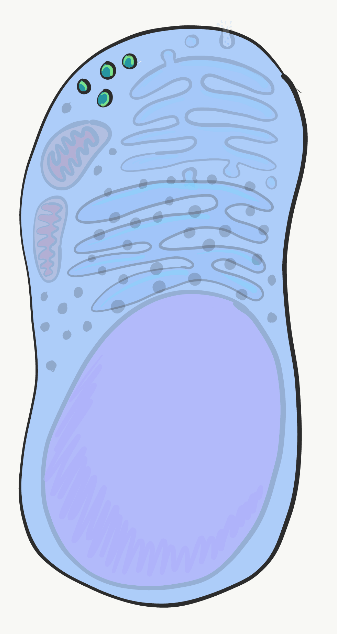
Cada célula humana está rodeada por la membrana plasmática. La membrana plasmática separa la célula de su entorno, y permite que ciertos materiales entren y salgan de la célula. Los fosfolípidos y el colesterol forman una barrera que separa a la célula del ambiente externo, lo que a su vez nos permite apuntar a esa célula y llamarla cosa. Las proteínas transmembrana abarcan la membrana plasmática y regulan lo que entra o sale. Otras proteínas transmembrana reciben señales de otras células o del ambiente, y transmiten esa información al interior de la célula. Otros más median el apego a estructuras fuera de la célula. Otras proteínas integrales de membrana se localizan en la superficie externa o interna de la membrana plasmática, pero no abarcan todo el ancho de la bicapa fosfolipídica. Muchas clases de biología se centran fuertemente en el núcleo, una tendencia que fue amonestada por uno de los primeros afroamericanos en obtener un doctorado, el doctor Ernest Everett Just, en su libro Biología de la superficie celular. El Dr. Just era embriólogo, y en embriología las proteínas de la membrana plasmática, especialmente los receptores y las proteínas de unión, son muy importantes. El citoplasma es el relleno gelatinoso de una célula, y en ocasiones es referido a un Fluido Intracelular (ICF). El citoplasma incluye nutrientes y electrolitos absorbidos por el fluido que rodea todas las células. Además, el citoplasma incluye una serie de proteínas y glicoproteínas sintetizadas por la célula. Estas moléculas pueden tener otras funciones importantes, pero atraen agua de la ECF. El resultado final es que las células tienen un relleno gelatinoso, en lugar de uno acuoso. Este relleno gelatinoso se rellena con una serie de orgánulos, similar a la forma en que la ensalada de gelatina de mi abuela contenía uvas, pasas y otros alimentos o sustancias similares a los alimentos que de ninguna manera convirtieron esa gelatina en una ensalada real.
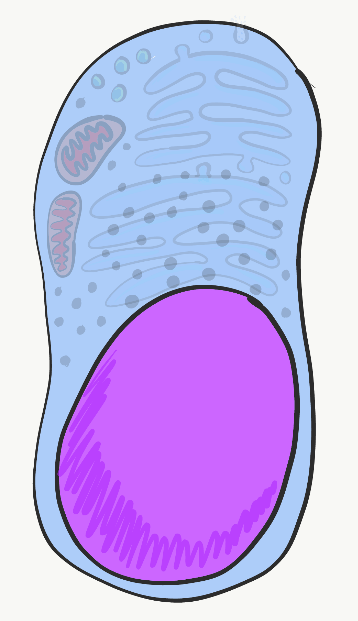
El núcleo
El núcleo contiene prácticamente todo el ácido desoxirribonucleico (ADN) de la célula. El ADN es las instrucciones para hacer una copia de ácido ribonucleico (ARN), y la mayor parte del ARN se traduce en el citoplasma en proteínas que se encuentran dentro y fuera de una célula. También son las instrucciones de cuándo y dónde elaborar estas proteínas. Por ejemplo, las células epiteliales de la mucosa oral no producen enzimas que secretan calcio y fosfato en la matriz extracelular. Por otro lado, las células epiteliales que se diferencian en ameloblastos elaboran estas proteínas, al expresar el ADN para estas enzimas, luego de que se lo hayan dicho otras células llamadas células de la cresta neural. Todas las células del cuerpo humano tienen el mismo ADN (con algunas excepciones). Sin embargo, diferentes células expresan diferentes ADN en diferentes momentos. El ADN se puede dividir en 2 tipos básicos. Hay genes, cada gen es más o menos las instrucciones para una sola proteína. El resto del ADN se pliega en formas únicas que proporcionan instrucciones sobre cuándo y dónde expresar estos genes. A este último se le conoce como ADN no codificante. Para que se produzca la expresión del ADN, las proteínas llamadas factores de transcripción (con la ayuda de moléculas pequeñas especiales) se unen a regiones reguladoras del ADN, abren la región apropiada y reclutan enzimas que copian una cadena de ADN en ARN mensajero (ARNm), que deja que el núcleo sea traducido en proteína. Otros factores de transcripción pueden inhibir temporalmente la expresión de genes.
Los factores de transcripción activan y apagan los genes rápidamente, en respuesta a los cambios en el entorno de una célula. Pero cuando las células se diferencian, cierran genes que no necesitan de manera más permanente. En lugar de depender de los interruptores de encendido y apagado de los factores de transcripción que se unen a las regiones reguladoras del ADN, estos genes inactivados se metilan (se une un grupo -CH 3), se empaquetan alrededor de proteínas histonas y se almacenan a largo plazo. El patrón de metilación del ADN y empaquetamiento de histonas se copia durante la mitosis. Esto significa que el patrón de genes que están disponibles o empaquetados es heredado por ambas células hijas, pero debido a que esta herencia no es una diferencia en la secuencia de ADN, se le conoce como herencia epigenética. Los rasgos epigenéticos son más capaces de modificarse en respuesta a los cambios en el ambiente que los genes (ADN), y juegan un papel importante en la diferenciación celular y el destino celular, que se trata con más detalle en los capítulos 8-11. Tenemos 46 moléculas de ADN en el núcleo— son moléculas terriblemente largas, pero sólo 46 en número— 23 maternas y 23 paternas. Durante la mitosis, estas 46 moléculas se empaquetan firmemente en 46 cromosomas (veces 2). Este empaque implica el uso de histonas a lo largo de casi toda la longitud de una cadena de ADN, y permite que los cromosomas se vean bajo el microscopio óptico. El resto del tiempo, el ADN se desenrolla en su mayoría (las instrucciones innecesarias se enrollan alrededor de las histonas, el resto son libres de transcribirse), y llena el núcleo de una manera que no se ve muy emocionante. A eso lo llamamos cromatina de ADN, y funcionalmente es de lejos la más excitante de las dos formas.

Ribosomas
Visible a lo largo del citoplasma son pequeñas motas hechas de proteína y ARN llamadas ribosomas. Estas estructuras traducen instrucciones de ARNm que provienen del núcleo, lo que significa que las instrucciones de ARNm se utilizan para guiar el enlace de una serie de aminoácidos en una proteína larga. Grupos de tres nucleótidos de ARNm, llamados codones, instruyen al ribosoma qué aminoácido agregar a una proteína. Al igual que las regiones reguladoras del ADN, el ARN que se encuentra en el propio ribosoma, no en las instrucciones de ARNm sino en el ARNr ribosómico, es un tipo de ARN que se pliega en formas específicas que, junto con las proteínas ribosómicas, realizan la reacción química de la traducción. Los ribosomas de flotación libre en el citoplasma sintetizan proteínas que permanecen en el citoplasma, como la queratina o enzimas que median la apoptosis.
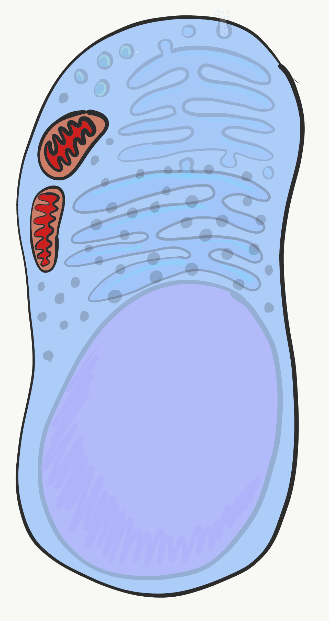
Mitocondrias
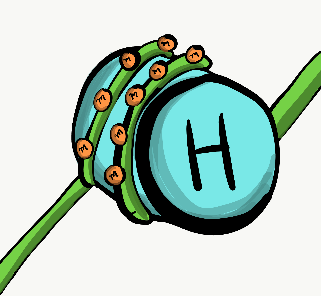
Las mitocondrias son donde se produce la mayoría de Adenosina Tri-Fosfato (ATP). El ATP está hecho de Adenosina, más tres grupos de fosfato (PO 4 3-), presta atención a la parte de fosfato, también es un componente importante del hueso, esmalte, dentina y cemento. El ATP potencia casi todos los procesos celulares, incluyendo la transcripción y traducción de proteínas mucosas dentro de una glándula salival, las señales eléctricas enviadas por las neuronas en la lengua cuando los alimentos ingresan a la cavidad oral y la contracción de las células mio-epiteliales para causar salivación. Las mitocondrias queman glucosa, usando oxígeno, y aprovechan parte de la energía liberada en forma de ATP. Las mitocondrias son diferentes de otros orgánulos en que contienen un poco de su propio ADN, que se hereda solo de la madre. Las mitocondrias también contienen dos membranas bicapa fosfolipídicas, no una como otros orgánulos. Utilizan esta membrana extra para generar ATP. Quizás cubriste la glucólisis y el ciclo del ácido cítrico antes. La parte a recordar es que las mitocondrias utilizan un gradiente de protones (o H +), lo que hace que el interior de las mitocondrias sea ácido, y por lo tanto potencialmente tóxico para el resto de la célula.
Lisosomas
Los lisosomas son pequeños compartimentos rodeados por los mismos fosfolípidos que en la membrana plasmática. Dentro de los lisosomas hay ácidos y enzimas digestivas que pueden usarse para destruir cosas dentro de la célula cuando se desgasta, o materiales que la célula ha engullido desde el exterior (por ejemplo, escombros, bacterias). Cuando una célula muere y comienza a romperse, las células vecinas están en peligro de ser dañadas por los ácidos y enzimas que se encuentran dentro de los lisosomas. En la cavidad oral, las células epiteliales solo tienen una vida útil de días antes de que se desgasten, por lo que es muy importante que estas células neutralicen primero los ácidos y enzimas en sus lisosomas, en un proceso llamado apoptosis.

Retículo endoplásmico
El Retículo Endoplásmico (RE) es una serie de tubos interconectados rodeados por una bicapa de fosfolípidos, similar a los lisosomas, solo más grandes, más tubulares y no llenos de ácido. El retículo endoplásmico liso (SeR) es donde las células producen lípidos y almacenan calcio. El retículo endoplásmico rugoso (RER) está cubierto de ribosomas. Las proteínas producidas por estos ribosomas terminan dentro del RER, luego viajan al aparato de Golgi y terminan siendo secretadas (como las proteínas mucosas) o permanecen dentro de la membrana plasmática (como las proteínas de unión celular, o receptores de moléculas de morfógeno).

Aparato de Golgi
El aparato de Golgi es otro conjunto de tubos, similar al RER. Pequeñas esferas encerradas en la membrana llamadas vesículas transportan proteínas elaboradas en el RER al aparato de Golgi, donde se modifican las proteínas. A menudo, estas proteínas tienen azúcares unidos a ellas, convirtiéndolas en glicoproteínas. Las vesículas llevan estas proteínas a la membrana plasmática, donde se secretan o se convierten en parte de la membrana plasmática. Cubrimos el papel que juega el colágeno proteico secretado en el esmalte y el ligamento periodontal. También cubrimos el papel compartido de la glicoproteína secretada fibronectina y la proteína unida a la membrana integrina tienen en la cicatrización del tejido gingival dañado.

Citoesqueleto
El citoesqueleto es una red de proteínas estructurales dentro del citoplasma. Esta red le da a la célula su forma, la capacidad de cambiar su forma, o de migrar. Se muestra en la Fig 1.10, las células tienen microtúbulos y filamentos de actina teñidos de rojo y verde, respectivamente. Estas proteínas no son generalmente visibles en imágenes histológicas más anticuadas, ambas serían rosadas en una tinción de H&E, al igual que las proteínas solubles en el citoplasma.
Fuera de la celda
| Matriz extracelular: |
|---|
| Sustancia molida |
| Fibras |
Cuadro 1.1: Dos componentes principales de la ECM. La matriz extracelular (ECM) incluye todo el material que se encuentra fuera de las células. Por lo general, se desglosa en los dos componentes enumerados en la Tabla 1.1.
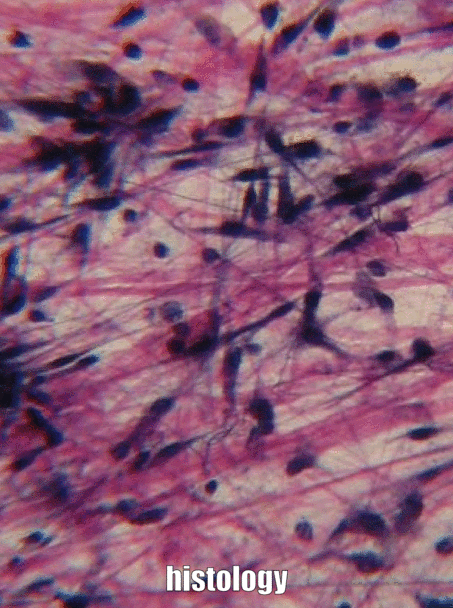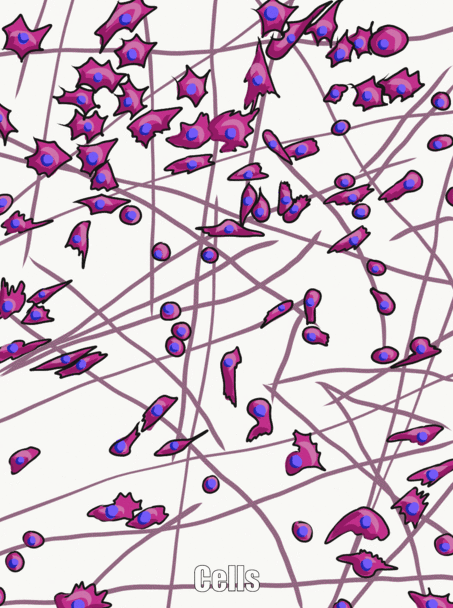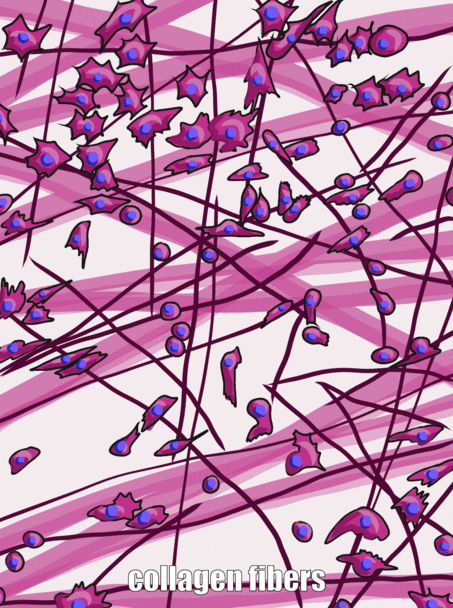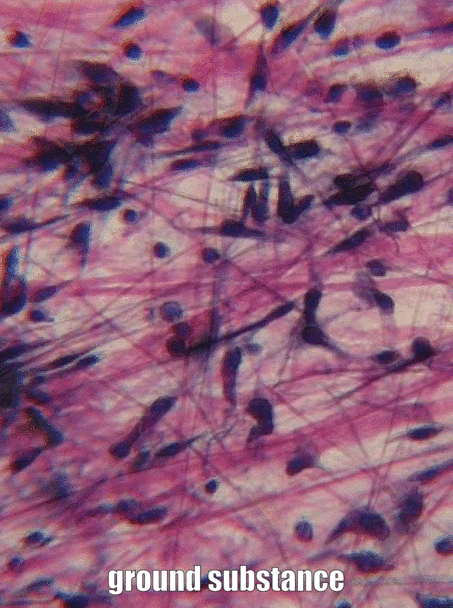
Sustancia molida
Sustancia molida es el nombre de todo el material fuera de la celda que no tiene una forma particular cuando se ve bajo microscopía tradicional. Una parte de la sustancia molida es el Fluido Extracelular (ECF), que es el agua y los nutrientes que rodean a las células que se llamaban plasma cuando estaba dentro de un vaso sanguíneo. Una vez que el líquido sale de la sangre y rodea las células, se llama ECF. A diferencia del plasma, este fluido es mantenido en su lugar por la otra parte importante de la sustancia molida, que incluye proteínas, glicoproteínas y polisacáridos. Estas moléculas son solutos, y su tamaño las hace inamovibles, por lo tanto son buenas para mantener el fluido en su lugar, formando un gel. No parece mucho bajo un microscopio, no más que si miraras muy de cerca alguna gelatina. Estas proteínas, glicoproteínas y polisacáridos son elaboradas y secretadas por las células (estas células probablemente tienen mucho RER y aparato de Golgi). Una de las glicoproteínas que se encuentran en la sustancia molida es la fibronectina, que es una proteína muy larga. Las células pueden reconocer, unirse y moverse a lo largo de la fibronectina si tienen la proteína integrina correcta que abarca su membrana plasmática. Por lo tanto, la fibronectina actúa no sólo como un camino por el que transitan las células, también es una hoja de ruta. Llegar las células adecuadas al lugar correcto en el momento adecuado es muy importante tanto en la curación como en el desarrollo. De hecho, lo que aprendes sobre el desarrollo se reutiliza (recapitula) en la curación. Otra molécula importante que se encuentra en la sustancia molida es un polisacárido grande llamado Ácido Hialurónico (HA). Al igual que la fibronectina, las células pueden unirse y viajar sobre HA (usando un tipo diferente de proteína de membrana plasmática), que tiene aplicaciones en odontología, como ayudar a las células de la encía a adherirse a un implante dental y formar un sello resistente a bacterias. No podemos ver la fibronectina o HA sin usar algunos trucos modernos de imagen, razón por la cual se enumeran como sustancia molida, y no en la siguiente sección, fibras. Debido a que las células migran sobre las proteínas de la sustancia molida, decimos que las proteínas ECM funcionan como un andamio. Sin andamios, los tejidos crecen solo desde sus bordes. Esto está bien si la velocidad no es importante, como cuando se están formando esmalte y dentina. Sin embargo, en la reparación de heridas, es óptimo que una herida se cure en todas partes a la vez, en lugar de desde los bordes. El cuerpo, por lo tanto, suele dejar primero algún tipo de andamiaje, como una costra. En odontología, se pueden crear andamios artificiales para ayudar al cuerpo a sanar, con base en nuestro conocimiento de las funciones de la sustancia fundamental. Ejemplos de andamios artificiales incluyen algunos materiales de injerto óseo, membranas periodontales y materiales. Además de su papel estructural como andamio, guiando las células a nuevas ubicaciones, las moléculas de sustancias terrestres proporcionan información a las células. Esta información le dice a las celdas dónde se encuentran, y qué deben estar haciendo. Por ejemplo, cuando una célula madre se une a la fibronectina, la fibronectina puede instruir a la célula madre para que exprese diferentes genes y se diferencie en un nuevo tipo de célula, como un odontoblasto, y comience a secretar dentina. Llegar las celdas a la ubicación correcta es agradable, pero necesitan saber qué hacer cuando lleguen allí. Esto ocurre durante la formación del diente y en respuesta a una lesión dental. A medida que aprendemos más sobre cómo la sustancia molida instruye a las células madre, mejoramos ayudando a los dientes a repararse ellos mismos. Es lamentable que muchos libros de texto pasen por alto la sustancia molida como solo el material gelatinoso fuera de una célula, razón por la cual hemos entrado en ella con más detalle, y en su lugar pasado por alto las mitocondrias.
Fibras
Tres proteínas extracelulares fueron visibles bajo un microscopio óptico hace un siglo, y se agruparon como fibras de la matriz extracelular. Al igual que la fibronectina y otras proteínas de sustancias del suelo, las fibras son secretadas por células llamadas fibroblastos.
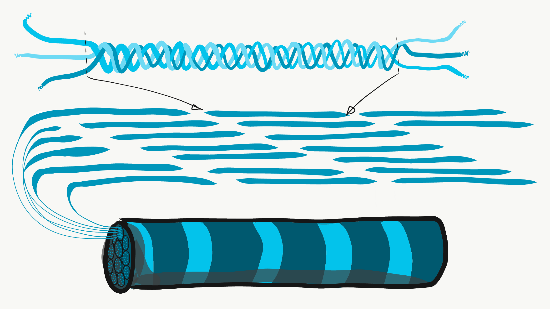
El colágeno es la más fuerte de las tres fibras y, por lo tanto, a menudo se le conoce como una proteína estructural, da forma a muchos órganos. Esta fibra está hecha de 3 hélices α-enrolladas, que a su vez están enrolladas y reticuladas entre sí, haciendo una macromolécula muy fuerte con la misma forma básica que una cuerda. Es muy fuerte si lo tiras de los extremos, pero se dobla si aplicas fuerza desde un lado. Por ejemplo, un área de caries avanzada en un diente puede ser tan suave como la madera flotante húmeda, ya que contiene sus fibras de colágeno sin los abundantes minerales que solían rodearlos. Las fibras de colágeno se encuentran en regiones de la cavidad oral donde es importante la capacidad de resistir la fuerza, como en el esmalte o el ligamento periodontal. De hecho, se encuentra en todo el cuerpo humano, representando el 25% de nuestro contenido proteico. Además de tener un papel estructural, el colágeno también actúa como andamio, similar a la fibronectina y HA, lo que permite que las células migren a través de las fibras largas. Obtener colágeno del RER (donde se traduce) al cuerpo de Golgi para su modificación, y finalmente secretado de la célula requiere vesículas extragrandes para su transporte. Las fibras reticulares, que no se muestran en la Figura 1.11, parecen una fina malla similar a una tela de araña bajo el microscopio. Posteriormente se descubrió que las fibras reticulares son diferentes formas de colágeno, pero todavía se les llama por su propio nombre, y muchas veces reciben igualdad de condiciones con el colágeno en la lista de fibras de ECM. Esta red de proteínas parecida a una red no es tan fuerte, pero proporciona suficiente marco para que las células sanguíneas descansen a lo largo de órganos como el bazo y los ganglios linfáticos, en lugar de hundirse hasta el fondo. Las fibras elásticas son más delgadas que las fibras de colágeno y, a menudo, se ven como pelos finos en el ECM bajo un microscopio. Están elaborados a partir de un tipo diferente de proteína (elastina). Como su nombre indica, estas fibras se pueden estirar y luego volver a su longitud original. Esto no es algo que el colágeno pueda hacer (si no puedes poner las palmas en el suelo mientras mantienes las piernas rectas, las fibras de colágeno de tus isquiotibiales y músculos gastrocnemios y sus tendones limitan tu rango de movimiento. El estiramiento regular puede activar la actividad de los fibroblastos y alargarlos). Las fibras elásticas se encuentran en mayor cantidad en regiones de la cavidad oral que cambian de forma durante el habla o la deglución, como el paladar blando.
Ni dentro ni fuera:
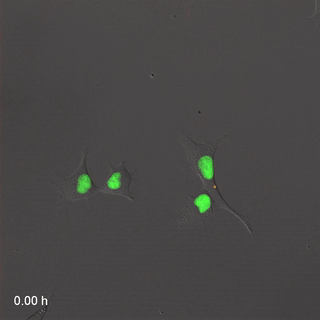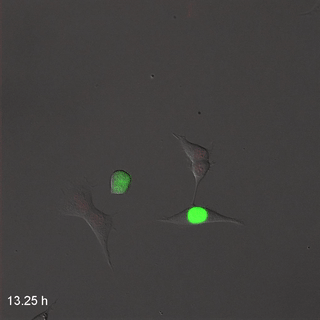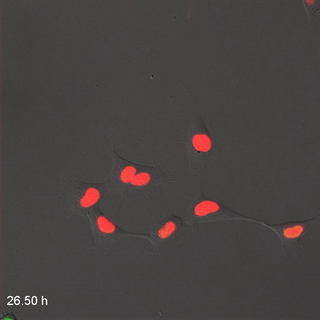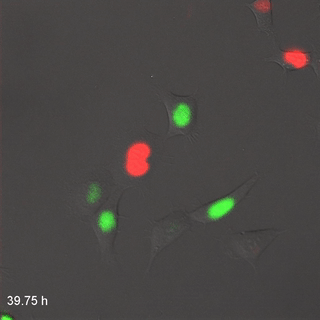
Mitosis
División celular, o mitosis, proceso por el cual una célula hace una copia de sí misma, produciendo dos células hijas idénticas. Temprano en el desarrollo, ya que estamos creciendo de una sola célula a un billón de células, se produce mucha mitosis. Cuando una célula no está experimentando mitosis, se dice que está en interfase. Este es el momento en que una célula podría estar haciendo su trabajo, como producir fibras para una nueva ECM, o una célula podría estar preparándose para la mitosis. Antes de que pueda ocurrir la división celular, una célula debe tener aproximadamente el doble de todo. Durante la mitosis, todo se divide a la mitad entre dos nuevas células hijas. No todas las células son capaces de la mitosis— de hecho, la mayoría de las células en un adulto se han diferenciado y están realizando tareas, están demasiado ocupadas para reproducirse. Decimos que estas células terminalmente diferenciadas han salido del ciclo celular. Para reparar el daño, los tejidos tienen células madre. Las células madre son células indiferenciadas que son capaces de dividirse y diferenciarse en uno o más tipos diferentes de células. Una célula madre se divide en dos células hijas, una hija normalmente sigue siendo una célula madre y la otra se diferencia en cualquier tipo de célula a la que se le indique. Por lo tanto, un tejido puede tener un suministro constante de células madre, siempre y cuando las células madre no mueran antes de sufrir mitosis. A medida que envejecemos, nuestros tejidos no sanan tan bien porque hemos perdido células madre. En términos generales, cuando una célula madre muere se va, otra célula madre no se somete a mitosis para producir dos células madre y reemplazarla. Las células madre se nombran en función de cuántos tipos diferentes de células pueden llegar a ser potencialmente. Las células madre unipotentes del epitelio oral se convierten en queratinocitos, y solo queratinocitos. Las células madre neuromesenquimales multipotentes se convierten en dentina, pulpa, cemento y ligamento periodontal. El óvulo fertilizado omnipotente se convierte en cada célula de un ser humano, además de más.

Para pasar por interfase y prepararse para otra ronda de mitosis, las células pasan por una serie de puntos de control del ciclo celular. Se trata de reacciones catalizadas por enzimas que impiden la progresión a la siguiente fase hasta que se acumula cierta cantidad de un producto Esto establece la tasa base para la división celular, lo que ayuda a asegurar que se produzca la cantidad correcta de crecimiento tisular. El paso a través de los puntos de control del ciclo celular está regulado por la fosforilación de proteínas llamadas ciclinas. Las ciclinas son factores de transcripción que activan genes que permiten la progresión a través del punto de control. La velocidad de este proceso puede ser acelerada o ralentizada por señales externas, como factores de crecimiento. Los factores de crecimiento son hormonas que se secretan en la sustancia fundamental de un tejido. La densidad y la adherencia de la sustancia del suelo influye en hasta qué punto se difunde el factor de crecimiento. Si la difusión es limitada, el factor de crecimiento solo acelera el crecimiento en un área localizada. Esto es importante en la formación de nuevos órganos, como los dientes. Otros factores de crecimiento podrían extenderse por un área amplia, especialmente si se secretan en el torrente sanguíneo. La secreción de Hormona de Crecimiento en la sangre, por ejemplo, permite que diferentes órganos crezcan aproximadamente a la misma velocidad. La mutación en un gen para un factor de crecimiento, o el receptor para un factor de crecimiento, puede conducir a una ganancia de función. Si las células reciben una señal de encendido constante para el paso a través del ciclo celular como resultado, llamamos a estos genes oncogenes. Solo se necesita una copia mutada de un gen receptor para obtener una función. Las ciclinas y otros genes que regulan el ciclo celular, sin embargo, se denominan genes supresores de tumores. Para que una célula pierda su capacidad de regular los puntos de control del ciclo celular, ambas copias de un gen supresor de tumores deben mutarse. En tu auto, tomaría un pie pisar el acelerador demasiado fuerte, pero tendrías que faltar ambos pies para no poder golpear los frenos. Por lo general, los cánceres se forman cuando una sola célula adquiere mutaciones tanto en oncogenes como en pares de alelos de genes supresores de tumores. Un alelo es una de las dos copias de un gen que se encuentran en la misma posición en dos cromosomas homólogos. Aunque esto ocurra, hay otra capa de protección cubierta es la siguiente sección.

Apoptosis
Todas las células contienen un grupo de proteínas receptoras de la superficie celular y enzimas intracelulares que les permiten sufrir muerte celular programada, o apoptosis, cuando se les instruye. La muerte celular programada es crítica para la vida multicelular, lo cual es algo extraño de decir. Sin ella, nuestros cuerpos estarían llenos de células muy viejas, que no funcionan. O peor aún, a medida que las células llegan al final de su vida —que para las células epiteliales y sanguíneas es muy corta— liberan el contenido de sus lisosomas y mitocondrias. Como recordarás, los contenidos de estos orgánulos son altamente ácidos, los cuales dañan o matan a las células vecinas. Si esas células vecinas murieran como resultado, ellas también liberarían su contenido lisosómico y mitocondrial, causando aún más daño. Cuando esto sucede en el cuerpo humano se llama necrosis tisular. Eso no es todo, una célula muerta arroja ADN, y el ADN es realmente largo y fibroso y tiende a ser muy pegajoso. Esto puede atrapar otras células y evitar que migren adecuadamente. Esto es genial si es genial si una célula está atrapando y matando bacterias y no le importa morir en el proceso, pero no es algo bueno que hacerle a las células humanas vecinas. Por lo tanto, se instruye a las células a sufrir apoptosis ya que están llegando al final de su vida útil, si el sistema inmunitario las ha determinado que están infectadas o cancerosas, o simplemente ya no son necesarias. La apoptosis asegura que antes de que una célula muera, neutraliza el pH de sus lisosomas y mitocondrias, y corta su ADN en pequeños trozos seguros. Durante el desarrollo, se producen más células de las necesarias, y las celdas adicionales se eliminan más tarde de manera organizada, similar a la forma en que la construcción de un edificio grande implica construir andamios primero, y el andamio se retira hacia el final del proyecto. El proceso de reparación de heridas también implica una sobreproducción de células seguida de su extracción organizada. Ojalá esto te parezca lógico para cuando termines este curso. Durante la reparación de heridas, se activan las instrucciones de ADN que fueron utilizadas por las células cuando se desarrollaban embriónicamente. Si quieres sonar elegante, y no todos nosotros, puedes decir que la cicatrización de heridas recapitula el desarrollo embrionario. Recapitular significa declarar de nuevo (repetir). El proceso de apoptosis comienza con una señal interna o externa. Por ejemplo, cuando el ADN de una célula se vuelve demasiado mutado, o si hay un número impar de cromosomas durante la mitosis, esto desencadena la apoptosis. Alternativamente, se puede instruir a una célula para que sufra apoptosis a partir de una señal extracelular llamada Factor de Necrosis Tumoral (TNF). Se activan una serie de enzimas llamadas caspasas, lo que finalmente conduce a la neutralización de los ácidos, la destrucción del ADN, y hacen que la célula explote en numerosos pequeños trozos, que pueden ser limpiados por macrófagos.
Uniones de célula a célula
| Tipos de uniones celulares | Ejemplos |
|---|---|
| Celda a celda | Desmosomas, uniones estrechas, uniones de hueco |
| Celda a ECM | Hemi-desmosomas |
Cuadro 1.2: Principales tipos de uniones celulares. Las uniones son grupos especializados de proteínas sobre o cerca de la superficie celular que hacen conexiones con alguna otra estructura. Estas conexiones pueden ser a otras celdas, o al ECM, como se indica en la Tabla 1.2.

Desmosomas
Las uniones de anclaje (desmosomas) son conexiones fuertes entre dos células. Las uniones de anclaje se emparejan y anclan el citoesqueleto de una célula a su vecina. Un gran grupo de células ancladas juntas por estas uniones son mucho más fuertes como grupo. “Una sola ramita se rompe, pero el haz de ramitas es fuerte” —Tecumseh. Los hemi-desmosomas son la mitad de un desmosoma anclado a la MEC, como el sello entre el epitelio gingival y la superficie no celular de un diente. Una de las muchas proteínas en un desmosoma es una integrina. Esta proteína reconoce y se une a proteínas en la ECM como la fibronectina. Cuando la proteína integrina de una célula se conecta a la fibronectina, esto no solo ancla el citoesqueleto de la célula a la ECM y ancla la célula en su lugar, la integrina también señala al interior de la célula, permitiendo que el núcleo sepa en qué tipo de tejido se encuentra la célula. Antes de que una celda pueda migrar a una nueva ubicación, primero debe eliminar sus uniones de anclaje. Durante el desarrollo, las células migran a nuevas ubicaciones y forman nuevas estructuras. Durante la cicatrización de heridas, las células madre se desprenden de sus vecinos, migran al área lesionada y comienzan la división celular para crear suficientes células para reparar la lesión.

Cruces apretados
Los cruces estrechos son uniones más pequeñas entre celdas. Las uniones estrechas rodean completamente una celda y crean un sello hermético entre esa celda y otra celda. Esto sirve para crear barreras entre una parte del cuerpo y otra, permitiendo que las células regulen lo que atraviesa y lo que no. Esto también le da a las células polaridad apical a basolateral (o una diferencia entre la parte superior e inferior), lo que es especialmente importante para un epitelio. El lado apical de una célula epitelial se enfrenta a la luz (centro hueco) de un órgano, mientras que el lado basolateral está más cercano al tejido conectivo subyacente. Las proteínas sintetizadas en el RER pueden enviarse a la porción apical o basolateral de la membrana plasmática. Una vez allí, las proteínas transmembrana del lado apical de la célula no pueden difundirse a la membrana plasmática en el lado basolateral, debido a que el anillo de uniones estrechas bloquea su movimiento.
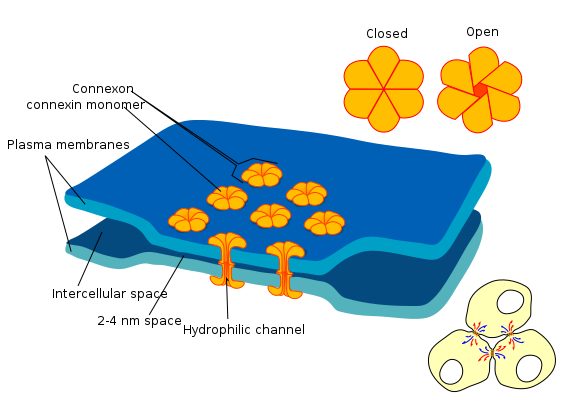
Cruces de separación
Las uniones gap (o conexones) son un grupo de proteínas que forman un pequeño paso entre las células, y pueden abrirse o cerrarse. Esta brecha permite que las células se comuniquen directamente entre sí. Debido a la forma en que las células epiteliales están conectadas entre sí, en una hoja, esta comunicación ocurre a través de un plano. Esta es una forma en que las células conocen su posición relativa a uno de los ejes del cuerpo, y es un proceso llamado Polaridad Celular Planar (PCP) (más lectura se puede encontrar aquí). PCP es polaridad de lado a lado, mientras que la polaridad apical a basolateral es de arriba a abajo. El PCP permite a las células saber en qué dirección están orientadas en el cuerpo, asegurando que las estructuras que están formando no solo estén en la ubicación correcta, sino en la orientación correcta. Por ejemplo, este tipo de señalización permite que los dientes se formen de manera que el lado lingual de un diente esté orientado hacia el lado lingual, no hacia los lados bucal, mesial o distal. La polaridad celular plana también ayuda a que los dientes erupcionen directamente en la cavidad oral, en lugar de hacerlo en ángulo.
Capítulo 0 * Capítulo 2