
1.2: Revisión Histológica
- Page ID
- 121782

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
- 4 tipos principales de tejido (por apariencia adulta)
- epitelios
- tejido conectivo
- músculo
- nervioso
- 3 tipos principales de tejidos (por linaje embrionario)
- ectodermo
- mesodermo
- endodermo

Una breve historia de histología
El objetivo de este capítulo es revisar la histología cubierta en tus clases de Anatomía y Fisiología, pero solo los tejidos que aparecen en la cabeza y el cuello, y solo las partes que son relevantes para este curso. Esto no es de ninguna manera una revisión integral. Si necesitas una revisión adicional sobre histología básica, prueba estos enlaces:
- Un diagrama de flujo interactivo de histología
- por Laird C. Sheldahl, Ph.D.
- El Laboratorio de Patología de Internet para la Educación Médica
- en la Universidad de Utah, Biblioteca de Ciencias de la Salud Eccles.
- HistologyGuide.com
- por T. Clark Brelje, Ph.D. y Robert L. Sorenson, Universidad de Minnesota.
La histología es el estudio de cómo se ven los tejidos bajo un microscopio. Un tejido es un grupo de células, todas del mismo tipo principal, que trabajan juntas para realizar una función. Debido a que la mayoría de las células son transparentes, los microscopistas utilizan una serie de tinciones diferentes para resaltar diferentes partes de las células. La Fig 2.1 es una imagen de una técnica de tinción muy común utilizando los colorantes Hematoxilina y Eosina (H&E). Estas dos manchas hacen que las moléculas con cargas negativas sean azules, y las cargas positivas de color rosa. El ADN y el ARN ocupan ambos colores, lo que hace que los núcleos sean púrpuras, mientras que la mayoría de las proteínas Desafortunadamente, la mayoría de las células están llenas de proteínas y su citoplasma se vuelve rosa, pero la matriz extracelular también está llena de proteínas, que también se vuelve rosa. Por lo tanto, a menudo estamos tratando de darle sentido a un mar de rosas con manchas moradas. En el lado positivo, si compras un libro para colorear de histología, solo necesitas dos crayones.

Imagenología
La mayoría de las imágenes que ves en los libros de texto de histología usan manchas como las manchas H&E, este tipo de tecnología se remonta al siglo XIX. La tecnología del microscopio ha mejorado desde entonces, como la imagen de la Fig. 2.2, que proviene de un microscopio confocal. El microscopio confocal utiliza costosos láseres para generar imágenes muy hermosas, pero se basa en manchas que teóricamente son similares a H&E. Lamentablemente, la mayoría de los libros de texto todavía usan imágenes anticuadas (más feas). Como no deberías estar usando ni los microscopios más nuevos ni los más antiguos en tu línea de trabajo, hemos ilustrado la mayoría de las imágenes histológicas en este libro. Esto nos permite enfocarnos en los conceptos, y dedicar menos tiempo a desarrollar las habilidades necesarias para interpretar imágenes de manchas rosadas con manchas moradas. Presumimos que en las clases de Anatomía y Fisiología que tomaste para ingresar a este programa, aprendiste a detectar las diferencias entre diferentes tipos de tejidos. Revisamos esas diferencias en este capítulo. Te pedimos que estés atento a las similitudes entre diferentes tejidos (aparte de que todos parecen manchas rosadas con manchas moradas). Si saltas adelante y lees los capítulos de embriología, tendrás una mejor idea de qué buscar, pero por ahora solo mantén ese pensamiento en la parte posterior de tu cabeza mientras cubrimos los principales tejidos del cuerpo humano.
Embriología suficiente para comenzar a aprender sobre histología

Clasificación: a la vieja manera, y una mejor manera.
La histología categoriza los tejidos en función de cómo se ven las células en la edad adulta. Si determinamos las familias de las personas en función de cómo se ven como adultos, podríamos hacer un buen trabajo al respecto, o podríamos cometer errores. Por ejemplo, podríamos agrupar las caras de dibujos animados en la Fig. 2.3 en dos familias basadas principalmente en el color del cabello y los ojos. Es así como se categorizan los 4 tipos principales de tejidos que vamos a revisar. El principal problema con este tipo de sistema de clasificación es que es difícil saber qué rasgos físicos se deben a la naturaleza (genética, como el color de ojos) versus la crianza (factores ambientales, como el tinte para el cabello).  Figura 2.4: Otra forma de categorizar rostros de dibujos animados, basados en datos de linaje.
Figura 2.4: Otra forma de categorizar rostros de dibujos animados, basados en datos de linaje.
Cuando aprendemos sobre embriología, sin embargo, vemos que podemos hacerlo mejor. Podemos categorizar los tejidos usando linaje celular en su lugar, rastreando con quién están relacionadas las células adultas siguiendo de qué célula provienen durante la embriogénesis. Los primeros embriólogos tiñeron células individuales y las siguieron a través de la división celular después de la división celular para ver en qué se convirtieron. Otros, como Hans Spemann, trasladaron celdas de un lugar a otro y preguntaron si todavía se convirtieron en lo que se esperaba, con base en su nueva ubicación. Al trasplantar solo unas pocas células conducen a embriones con dos cabezas completas, Spemann concluyó que fue inesperado. Para tener una mejor idea de cómo sucede eso, la ciencia tuvo que esperar el descubrimiento de información genética que almacena ADN para un organismo. Entonces, biólogos del desarrollo como Christiane Nüsslein-Volhard pudieron determinar los mecanismos por los cuales una sola célula embrionaria se convirtió en diferentes tipos de células en un organismo adulto, su organismo preferido eran las moscas, y por ese trabajo ganó el Premio Nobel. A diferencia de los histólogos y embriólogos anteriores que nombraron a todo después de ellos mismos, obtenemos nombres divertidos como Bicoid, Dickkopf, Frizzled y Sonic Hedgehog. Estos científicos siguieron los linajes de las células, observaron en qué se convierten y determinaron cómo las células toman decisiones sobre cuándo y en qué convertirse. En lugar de usar la apariencia de nuestra gente de dibujos animados, si conociéramos su linaje (también podrías ver el término ontogenia más elegante), podemos clasificarlos en familias más precisas, con una subfamilia (Fig. 2.4). Cuando veas la palabra linaje, piensa en línea familiar, donde las células diferenciadas son la generación más joven, y las células madre son sus padres y abuelos. Tristemente, no es así como se enseña la histología, así que vamos a ver cómo son las células adultas y clasificarlas en función de las características físicas. Cuando lo hacemos, mira para ver si dos tejidos tienen bordes distintos (probablemente linajes diferentes) o bordes mezclados (a menudo el mismo linaje).
| Los 4 principales tipos de tejidos |
|---|
| epitelial |
| Tejido Conectivo |
| Músculo |
| Nervioso |
Cuadro 2.1: los 4 tipos principales de tejido
Epitelios
Ubicación del epitelio
Un epitelio está compuesto por células epiteliales, generalmente conectadas entre sí en una lámina. Hay muy poca MEC en un epitelio, en su mayoría células. Las células están ancladas juntas por desmosomas, los cuales las sujetan en una lámina. Las uniones estrechas aseguran que las cosas que viajan de un lado del epitelio al otro pasen a través de las células epiteliales, y por lo tanto deben ser solubles en lípidos, o ser reconocidas por las proteínas transportadoras en las membranas plasmáticas apicales y basolaterales. Por lo tanto, un epitelio tiene polaridad apical a basolateral. La superficie apical mira hacia el exterior del cuerpo, y el basolateral hacia adentro, y polaridad significa que los dos lados son diferentes. El exterior puede significar la superficie externa de la piel, o una superficie interna, como recubrir la cavidad oral, el estómago y la vejiga. Toda superficie del cuerpo humano es un epitelio. Por lo tanto, bajo el microscopio, si ves un espacio vacío, un epitelio bordea ese espacio. El revestimiento interno de un conducto sudorídico es un epitelio. El revestimiento interno de un vaso sanguíneo es un epitelio. Todos los demás tejidos se encuentran entre ese epitelio y el epitelio en la superficie externa de nuestro cuerpo. Incluso el esmalte está hecho por un epitelio, así que si alguien te dice que el esmalte es un tejido conectivo porque se ve similar al tejido óseo, no han estudiado embriología, y necesitan ser educados. Usa palabras como ontogenia.

Los epitelios son delgados
El tejido epitelial es bueno para la curación porque contiene numerosas células madre capaces de someterse a mitosis (se puede decir por la presencia de cromosomas visibles, en lugar de cromatina). Para epitelios más gruesos, las células madre residen en la capa más profunda. Los epitelios no pueden ser terriblemente gruesos, sin embargo, porque son avasculares (sin sus propios vasos sanguíneos). Los nutrientes para un epitelio deben difundirse desde el tejido conectivo subyacente, razón por la cual en la Fig 2.6, las células epiteliales del lado apical carecen de núcleos. Están muertos, pero aún conectados con el resto del epitelio. El inconveniente de ser bueno en la mitosis es que significa que esas células están unos pasos más cerca de volverse cancerosas. Muchas personas se exponen regularmente a altas dosis de carcinógenos que se encuentran en las bebidas alcohólicas y el tabaco (en particular el tabaco de mascar), lo que conduce a cánceres orales. Además de las membranas, las glándulas están hechas de células epiteliales. Las glándulas exocrinas, como las glándulas salivales, contienen una sola capa de células epiteliales, enrolladas en tubos llamados conductos. Estos conductos llevan secreciones exocrinas a una superficie del cuerpo. Las glándulas endocrinas, sin embargo, son diferentes. No están dispuestos en una lámina, sino que son racimos amorfos de células epiteliales. Las glándulas endocrinas tampoco son avasculares. Los vasos sanguíneos crecen directamente en el epitelio de una glándula endocrina. Esto es importante porque las glándulas endocrinas secretan hormonas directamente a la sangre.
| Número de capas | Forma de celdas |
|---|---|
| Sencillo | Escamoso |
| Estratificado | Cuboidal |
| Columnar |
Cuadro 2.2: La categorización básica de la mayoría de los epitelios.
Los epitelios se clasifican en su mayoría con base en dos criterios. Con algunas excepciones, los epitelios tienen un nombre de la primera columna y un nombre de la segunda columna del Cuadro 2.2. Los epitelios simples tienen solo una capa de células, mientras que los epitelios estratificados tienen más de una. Escamosas significa que las células son planas, como huevos fritos. Cuboidal significa cuadrado en apariencia (las células son 3D, pero se ven 2D bajo el microscopio). Columnar significa que las células son altas (como columnas).
 Figura 2.7: Epitelios escamosos simples (endotelios) que forman las paredes de los capilares.
Figura 2.7: Epitelios escamosos simples (endotelios) que forman las paredes de los capilares.
Epitelio escamoso simple
Si miras el revestimiento de un capilar, es posible que puedas ver el epitelio escamoso simple que recubre la superficie interna. Estas células, llamadas células endoteliales, son funcionalmente células realmente frías, aunque sean difíciles de ver. Normalmente, las células endoteliales mantienen la sangre dentro de un capilar, pero pueden dañarse, como durante el sondeo, el uso del hilo dental o la tos.

Epitelio cuboidal simple
Los epitelios cuboidales simples alinean la mayoría de los conductos de las glándulas exocrinas, como las glándulas sudoríparas y salivales. En la Fig 2.8, debería ver dos conductos saliendo hacia la cámara, y un conducto que discurre de lado a lado. El lumen de cada conducto está revestido por un simple epitelio cuboidal. Las células cuboidales en el epitelio están conectadas a sus vecinos en los lados mediante uniones de anclaje. Donde dos conductos se acercan, se trata de dos epitelios simples cuboidales, no uno estratificado, hay dos capas separadas de epitelios, con un poco de sustancia molida entre ellos. Existen epitelios cuboidales estratificados, como en partes de la glándula salival parótida, pero son lo suficientemente raros como para que podamos saltarlos. Los capilares de la Fig. 2.7 están revestidos por células planas para permitir la difusión de nutrientes fuera del capilar y los productos de desecho hacia adentro, mientras que los conductos salivales en la Fig. 2.8 tienen células más gruesas para evitar que la saliva se filtre y disuelva los tejidos conectivos de la cara.

Epitelio columnar simple
Los epitelios columnares simples no se encuentran en la cavidad oral adulta, pero en un embrión el epitelio del esmalte interno columnar se diferencia en ameloblastos productores de esmalte durante la embriogénesis. La forma de las células no es importante aparte de que ayuda a los histólogos a identificar estas células en un diente en desarrollo.
 Figura 2.10: Vista lateral de la cama, mostrando un gato entre la superficie basolateral del edredón y la superficie apical de la sábana delgada. Crédito de la imagen: “foto” por desconocido está en el Dominio Público, CC0
Figura 2.10: Vista lateral de la cama, mostrando un gato entre la superficie basolateral del edredón y la superficie apical de la sábana delgada. Crédito de la imagen: “foto” por desconocido está en el Dominio Público, CC0Si estás viendo la ilustración de la Fig 2.9, tal vez te preguntes por qué hay menos núcleos de dibujos animados que los visibles en la imagen histológica. ¿El ilustrador te dio por error una imagen de un epitelio columnar estratificado e intentó hacerla pasar por simple? No. Cuando decimos que un epitelio es simple, significa una célula gruesa de apical a basolateral. Pero cuando cortamos una muestra de tejido para verla bajo el microscopio, el corte puede tener varias células de un lado a otro. Y cuando colocamos la muestra de tejido de lado, las células que corrían de lado a lado ahora están sentadas una encima de la otra. Hemos ilustrado cómo debería ser un epitelio simple si tuviéramos la porción ideal de tejido de una célula gruesa. Este podría ser un buen momento para sacar a colación la perspectiva. Los epitelios forman láminas planas, y en la Fig. 2.9 estamos viendo una lámina (plegada) en sección transversal. Imagínese cortando su cama y mirándola desde la vista lateral. Podrías distinguir las capas individuales y individuales de tu sábana y edredón. Además, podrías notar que si bien cada una es de una sola capa, la sábana es delgada y el edredón es más grueso. Podrías identificar la superficie basolateral de tu sábana (el lado que te toca cuando duermes) y la superficie apical (el lado cercano al edredón). En la Fig. 2.10, si hubiera un segundo gato encima del primer gato, podríamos decir que las capas de gatos están estratificadas. Pero si la sábana estuviera cubierta en una sola capa de gatos, de lado a lado y de adelante hacia atrás, la capa de gatos sería simple. Bajo el microscopio, estamos haciendo lo mismo, pero desafortunadamente las células son semitransparentes, por lo que se vuelve desordenado. Cuando llamamos a esto un epitelio simple en la Fig. 2.9, eso significa que solo hay una célula rosada entre el espacio blanco (aquí, la luz de los intestinos) y el espacio muy azul claro (el tejido conectivo, donde los nutrientes pueden ser absorbidos en los vasos sanguíneos). Esas celdas son más gruesas que las que observamos en la Fig. 2.8, pero no hay columnas encima de otras columnas. 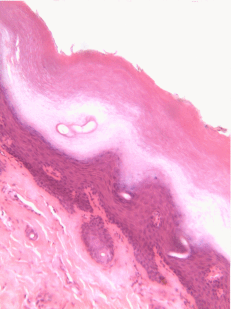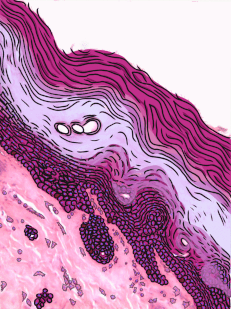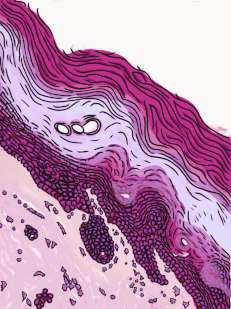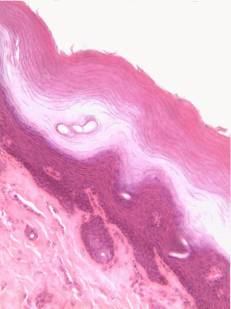
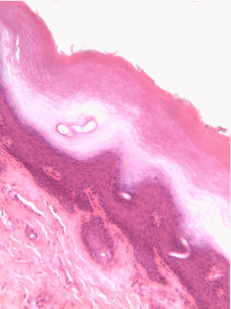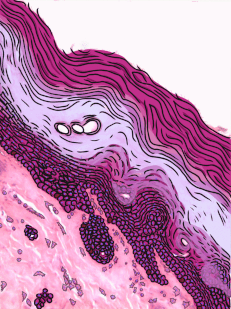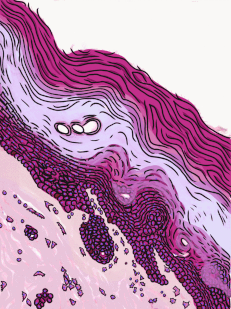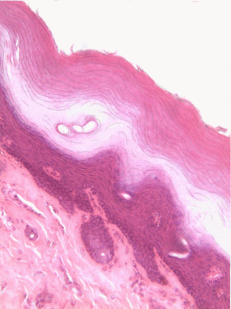
Figura 2.11: Histología y caricatura de un epitelio escamoso estratificado (y un poco de tejido conectivo debajo).
Epitelio escamoso estratificado
Los epitelios escamosos estratificados se encuentran en el revestimiento externo de la piel y en el revestimiento interno de la mucosa oral. Si bien tenemos diferentes nombres para la piel y la mucosa oral, si los nombráramos en base a la embriología, más que a la apariencia superficial, ambos contendrían un epitelio escamoso estratificado. El capítulo 3 abarca con más detalle la epidermis de la piel, que es un epitelio escamoso estratificado queratinizado. La queratina es una proteína resistente y resistente al agua elaborada por el principal tipo de célula epitelial en este tejido, el queratinocito. El epitelio escamoso estratificado cambia en el borde bermellón a menos queratinizado, y el epitelio escamoso estratificado de la mucosa oral es menor o no queratinizado. Con base en esa información, ¿debemos clasificar la zona bermellón como una extensión de la mucosa labial, o como parte de la piel y los labios? ¿O hay una línea en la zona bermellón que se divide en una mitad de piel y una mitad de mucosa oral? En momentos como este podríamos usar el consejo de un embriólogo para informarnos que no siempre debemos buscar líneas en los tejidos adultos, ni definir fronteras entre la piel, el bermellón y la mucosa oral en función del color. Si cambias de carrera e ingresas a proctología, usa esta misma información ahí abajo.
Epitelio estratificado cuboidal, columnar estratificado y transicional
Estos tejidos no aparecen en este libro de texto.

Epitelios pseudoestratificados
El epitelio columnar pseudoestratificado ciliado, o como dicen muchas personas, epitelio pseudoestratificado, se puede encontrar en las partes superiores del tracto respiratorio, como recubrir la cavidad nasal y los senos paranasales. Este tejido definitivamente merece un nombre diferente al epitelio escamoso estratificado de la piel, labios y mucosa oral. Sin embargo, probablemente no necesitó romper las reglas de nomenclatura descritas en la Tabla 2.2. Un epitelio pseudoestratificado tiene más de una capa, pero es difícil contar cuántas. En consecuencia, alguien llamó a este tejido pseudo estratificado. Las células azules grandes y gordas de la Fig. 2.12, que no tienen cilios, son células caliciformes. Estas células producen moco. Las células caliciformes sintetizan proteínas mucosas dentro de su RER y las secretan, y cuando atraen agua, se convierten en moco. Esto es muy similar a la sustancia molida, sin embargo el moco se secreta fuera del cuerpo, la sustancia molida es una parte del cuerpo. La coloración azul de las células caliciformes nos dice que las proteínas mucosas no portan las mismas cargas iónicas que la mayoría de las proteínas que se encuentran dentro de las células epiteliales del epitelio pseudoestratificado. Debido a esta diferencia de color, se clasifican como una célula distinta, una glándula unicelular que se encuentra dentro del epitelio pseudoestratificado. Pero, ¿qué dice nuestro amigo el embriólogo? Bueno, las células columnares no pueden convertirse en células caliciformes, y las células caliciformes no pueden convertirse en células columnares, pero sí comparten un ancestro común (o célula madre), por lo que probablemente deberíamos considerar a las células caliciformes como parte del epitelio pseudoestratificado. ¿Importa? No, pero preguntas como esta importan más adelante en el libro, es bueno empezar a practicar ahora.
Tejido conectivo (TC)
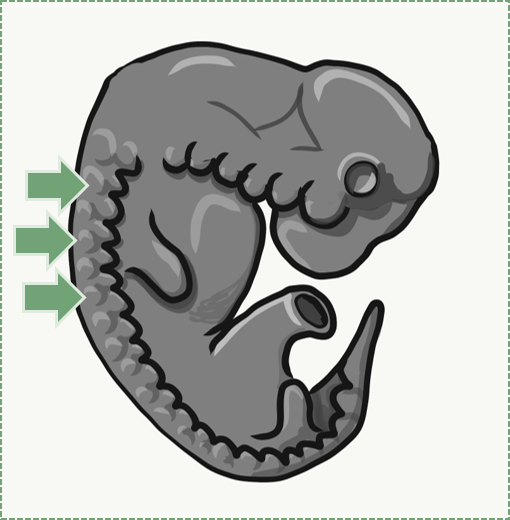
Características generales del tejido conectivo
Muchos libros de texto te dicen que el tejido conectivo conecta otros 2 tejidos. Eso es como decir que tu teléfono inteligente te conecta con otras personas. Es técnicamente cierto, pero pasa por alto el hecho de que estas podrían ser personas que conoces, personas que no conoces, o personas que no quieres conocer. Ignora el hecho de que podrías estar comunicándote sincrónicamente con personas del otro lado del mundo, comunicándote asincrónicamente por correo electrónico, accediendo a información del conocimiento humano casi suma-de-todo (internet), o leyendo la perorata de segundo año de algún arrastrador de nudillos descontento que, hace años, publicó un comentario grosero sobre una grabación en video en YouTube de una conferencia de clase de histología (también internet). Hasta el momento, hemos tratado de mantenerlo a usted, el lector, a salvo de información extraña sobre histología que no sea relevante para su carrera. Aquí entramos en más detalles que tu clase promedio de histología de pregrado. Los tejidos conectivos provienen de segmentos de tejido en un embrión, que se encuentran entre dos capas de epitelio, llamadas somitas (Fig. 2.13). Debido a su linaje compartido, los tejidos conectivos comparten una serie de características comunes. El tejido conectivo promedio tiene pocas células, y en su mayoría está compuesto por Matriz Extracelular. Esa matriz incluye glicoproteínas y polisacáridos elaborados por las células, que atraen el agua para formar la matriz de gel de sustancia molida. También incluye fibras más visibles. Algunas moléculas importantes de sustancia fundamental cubiertas anteriormente son Fibronectina y Ácido Hialurónico. Los tres tipos principales de fibras visibles son las fibras de colágeno, reticulares y elásticas. Los tejidos conectivos suelen ser altamente vasculares, lo que significa que contienen vasos sanguíneos. Hay varios tipos de células diferentes que se encuentran dentro de un tejido conectivo. Las células madre son células madre mesenquimales (MSC). Las células madre mesenquimales son células extremadamente importantes (aquí hay más lecturas), y como las puedes encontrar en tejidos adultos se les llama un tipo de célula madre adulta. Estas células son capaces de sufrir mitosis para producir más células madre adultas, las cuales pueden diferenciarse en fibroblastos, lipoblastos, condroblastos, osteoblastos, hemocitoblastos, mioblastos y neuroblastos. Así, forman la mayoría de los tejidos conectivos, incluyendo hueso, cartílago, sangre (tanto glóbulos rojos como blancos), más tejido muscular. Eso es prácticamente de todo excepto los epitelios, que las células madre mesenquimales también pueden formar después de pasar por una transición.

En un tejido conectivo básico, las células madre mesenquimales se dividen y se diferencian en fibroblastos, las células que explotan las fibras y la sustancia molida que se encuentran en la MEC. Se pueden encontrar otras células en un tejido conectivo, incluyendo adipocitos, glóbulos rojos y blancos, u otras células que hayan emigrado de su tejido de origen. En un tejido conectivo maduro, los fibroblastos a veces se llaman fibrocitos, lo que encaja con nuestra nomenclatura regular para una célula que atraviesa etapas de diferenciación. Con mayor frecuencia, sin embargo, se les llama fibroblastos y no nos preocupamos de si están volando activamente las fibras en su momento o tomando un descanso.

Mesénquima
El mesénquima es el primer tipo de tejido conectivo que produce un embrión. Se diferencia en otros tejidos conectivos. Se trata de un tejido embrionario que aún tiene que decidir en qué se convertirá cuando crezca. Está compuesto principalmente por células madre mesenquimales y sustancia mucosa molida (mucosa es un adjetivo para describir la MEC suelta, mientras que la mucosidad es una sustancia excretada del cuerpo). Existe un tipo especial de mesénquima importante para el desarrollo facial y dental, llamado neuromesénquima. El linaje del neuromesénquima no proviene del mesodermo como la mayoría de los tejidos conectivos, sino de un tipo especial de célula neuronal llamada célula de la cresta neural. El neuromesénquima forma tejidos conectivos especiales que aún no hemos mencionado: la dentina, el cemento y el ligamento periodontal.

Tejido conectivo areolar
El tejido conectivo areolar es el tejido conectivo por excelencia, o el más aburrido dependiendo de cómo lo mires. Contiene un poco de todo lo que otros tejidos conectivos podrían tener: células, sustancia fundamental y las 3 fibras. Debido a que tiene una buena cantidad de sustancia molida, es un tejido ideal para ocupar lugares donde los vasos sanguíneos podrían necesitar espacio para crecer en el futuro. De ahí que la TC areolar se encuentre en regiones que son altamente vasculares. Es por ello que se puede encontrar una pequeña capa de tejido conectivo areolar directamente debajo de casi todos los epitelios, incluyendo el epitelio escamoso estratificado de los diversos tipos de mucosa oral. La sustancia molida también absorbe agua y se hincha de tamaño, razón por la cual el tejido conectivo areolar de la cavidad nasal también se conoce como tejido eréctil.

Tejido conectivo irregular denso
Debajo de la TC areolar de la mucosa oral se encuentra tejido conectivo irregular denso. Los fibroblastos forman la mayor parte del tejido: las fibras de colágeno, las más fuertes de los 3 tipos de fibras. En una TC irregular densa, las fibras de colágeno apuntan en todas las direcciones. Esto hace que este tejido sea particularmente fuerte en todas las direcciones. En la Fig. 2.17, la mayor parte del rosa son fibras extracelulares de colágeno, y la sustancia molida del espacio blanco. Se puede encontrar TC irregular densa (junto con su compañero, TAC areolar) en la dermis de la piel. ¿Por qué en ambos lugares, preguntas? Linaje.
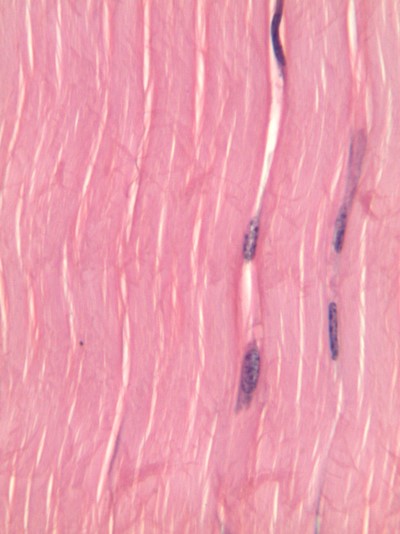
Tejido conectivo regular denso
Al igual que la TC irregular densa, el tejido conectivo regular denso es principalmente fibras de colágeno, solo las fibras corren paralelas entre sí. Eso hace que este tejido sea fuerte en una dirección. En su mayoría encuentras TC regular densa entre músculo y hueso, o entre hueso y hueso. El único lugar en este libro donde encontramos TC regular densa es el ligamento periodontal, entre hueso y cemento. Podrías estar pensando que las fibras de colágeno se ven onduladas, no paralelas. Cuando alguien cortó esta sección de un tendón, cortaron el tejido usando una cuchilla delgada. A medida que una cuchilla corta el tejido, el tejido se mueve, y así la cuchilla empuja las fibras en una dirección, luego las tira hacia atrás y empuja de nuevo a medida que la persona corta una rebanada lo más delgada que pueda. Para obtener mejores resultados, use un dispositivo costoso que congela o incruste tejidos blandos en cera, como un micrótomo, critoma o vibratoma. Traemos esto porque muchos libros de texto sugieren que la histología es una representación casi perfecta de lo que se encuentra en el cuerpo humano, cuando en realidad hay artefactos, o errores, que los histólogos han aprendido a ignorar cuando los vemos. Hemos optado por ilustrar muchas de las imágenes de este libro para evitar la discusión de artefactos donde sea posible, y enfocarnos en conceptos relevantes para la higiene dental.

Tejido conectivo adiposo
El tejido adiposo es un tipo de tejido conectivo. Es especial en que se tiene muy poco ECM. Contiene en su mayoría adipocitos, las células maduras que almacenan triglicéridos. También contiene células madre mesenquimales, que si una persona necesita más tejido adiposo, se dividen y se diferencian en más adipocitos. Técnicamente, las Células Madre Mesenquimales se diferencian primero en un lipoblast, pero esto solo es importante para mantener la coherencia con el sistema de nomenclatura. A lo mejor si hubieran nombrado a las células adipoblast y adipocito, usaríamos el nombre -blast con más frecuencia, pero alguien lo estropeó. Muchos libros de texto colocan el tejido adiposo junto al tejido conectivo areolar y los clasifican a ambos “sueltos”, debido a su bajo número de fibras visibles. Eso está bien si te gustan las categorías basadas en apariencias, pero no necesitas llamar a ninguna de ellas “suelta”. Es descriptivo y no corresponde a nada embriológico. Encontramos tejido adiposo en muchos lugares del cuerpo, ya sea entre otros dos tejidos, o incrustado dentro de otro tejido (como el adiposo que encuentras en el tocino). Por otro lado, el tejido conectivo areolar se encuentra junto al tejido conectivo irregular denso, razón por la cual nos hemos desviado de la agrupación que se encuentra en la mayoría de los libros de texto.

Tejido conectivo cartilaginoso
El tejido conectivo del cartílago viene en tres sabores básicos: cartílago hialino, fibrocartílago y cartílago elástico. La característica distintiva del tejido conectivo del cartílago es su amplia cantidad de sustancia molida. Los condroblastos secretan glicosaminoglicanos y glicoproteínas, como el Ácido Hialurónico, que atraen el agua, formando un gel muy denso pero resbaladizo. La MEC del cartílago contiene colágeno— hay más colágeno en el fibrocartílago de la articulación temporo-mandibular, mientras que el cartílago elástico de la epiglotis tiene más fibras elásticas. Después de secretar la MEC, los condroblastos se diferencian en condrocitos, y residen dentro del tejido en espacios llamados lagunas (lagos). Los bordes externos del cartílago son una fina capa de fibras de colágeno paralelas conocidas como pericondrio. Si nos fijamos en la Fig 2.20, el pericondrio aparece como una capa de tejido conectivo regular denso (lo es, solo tiene un segundo nombre), y el borde del pericondrio se funde en tejido conectivo denso irregular. La razón por la que el cartílago se mezcla con tejido conectivo irregular denso es que comparten el mismo linaje: los condroblastos y los fibroblastos se diferencian de las células madre mesenquimales.
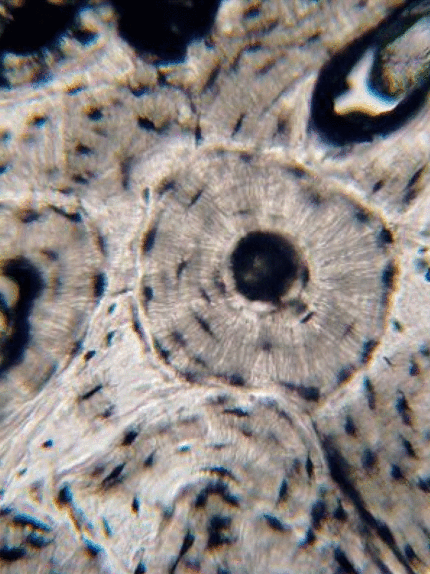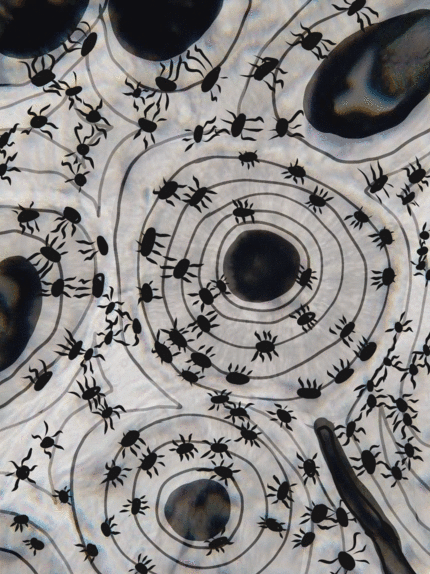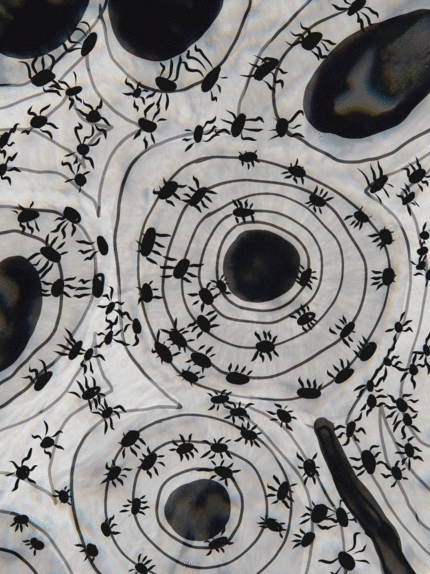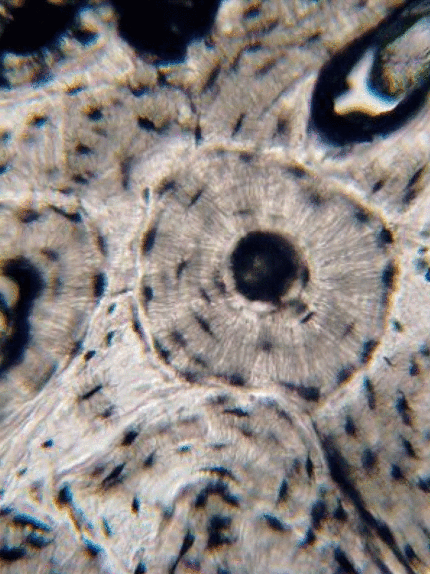
Tejido conectivo óseo
El tejido óseo (o tejido óseo) es el último tipo de tejido conectivo que cubrimos en detalle. Debido a su similitud con la dentina, el cemento y el esmalte, cubrimos el tejido óseo con bastante detalle. Como cualquier tejido conectivo, el tejido óseo comienza como células madre mesenquimales, que se diferencian en una célula madre intermedia llamada célula osteo-condroprogenitora, que a su vez puede decidir adoptar un destino de osteoblastos o condroblastos, dependiendo de qué señales reciba. Una célula madre que haya adoptado el destino de los osteoblastos (y negado el destino de los condroblastos) sería una célula osteo-progenitora. Estas células osteo-progenitoras son células madre dentro de los tejidos conectivos que rodean los huesos, el periostio (superficial) y el endosteo (profundo). Cuando se estimulan, se someten a mitosis, y una hija se diferencia en un osteoblasto. Un osteoblasto es la célula que secreta la ECM que conforma la mayor parte del tejido óseo. Los osteoblastos secretan fibras de colágeno y sustancia molida, que posteriormente mineraliza, atrapando los osteoblastos dentro de una laguna (un pequeño espacio dentro de la ECM densa). En este punto, se diferencian en osteocitos y mantienen el tejido óseo. A continuación, un nuevo conjunto de osteoblastos deposita otra capa de tejido óseo alrededor de la anterior. Esto crea las capas concéntricas de tejido óseo que se encuentran en los osteones del hueso compacto, o las capas espirales más delgadas de tejido óseo que se encuentran en las trabéculas del hueso esponjoso.
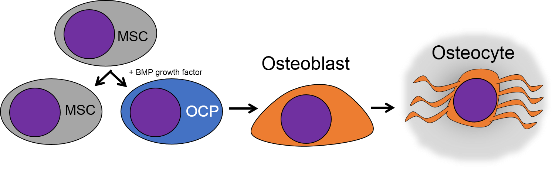

La matriz extracelular del tejido óseo es aproximadamente 1/3 fibras de colágeno (proteína orgánica) y 2/3 minerales (inorgánicas). El componente mineral es una mezcla de iones Ca 2+ cargados positivamente, los cuales reaccionan con fosfato de carga negativa (PO 4 3-) para formar una matriz cristalina dura. Debido a que la mayoría de las células están llenas de fosfato, los niveles de Ca 2+ en el citoplasma siempre deben mantenerse muy bajos, por lo que si una célula como un osteoblasto desea almacenar Ca 2+, lo hace dentro de su retículo endoplásmico liso. El Ca 2+ PO 4 3- reacciona adicionalmente con agua (o más específicamente, iones hidróxido OH —) y pequeñas cantidades de fluoruro (F —) para formar un cristal llamado hidroxiapatita de calcio. La dentina, el esmalte y el cemento tienen ECM muy similar al tejido óseo, solo con cantidades variables de colágeno versus componentes minerales. Las fibras de colágeno corren paralelas dentro de una capa de tejido óseo y 90° en la siguiente capa. Si bien el componente mineral de los tejidos óseos y dentales proporciona fuerza, el colágeno proporciona la capacidad de doblarse o flexionarse, reduciendo las posibilidades de que el tejido se corte bajo tensión. Las fibras de colágeno, por lo tanto, tienen una función similar a la de las barras de refuerzo en concreto armado. Los menores porcentajes de colágeno que se encuentran en el esmalte lo hacen más duro y resistente a la caries que la dentina o el cemento, pero aún susceptible a la fractura si se aplica suficiente fuerza.
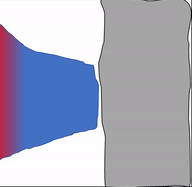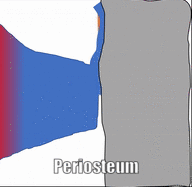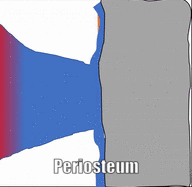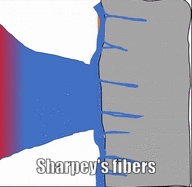
Los huesos están rodeados por una capa de denso tejido conectivo regular llamado periostio. Este tejido es una continuación (no una estructura separada) del mismo tejido de tendones y ligamentos. El periostio también contiene células osteo-progenitoras, las células semi-diferenciadas que provienen de las Células Madre Mesenquimales, así como osteoblastos y osteoclastos (subiendo). Los haces de fibras de colágeno de esta capa llamada fibras de Sharpey penetran en las capas superficiales del hueso compacto. Esto hace una conexión muy fuerte entre el tejido óseo y los tendones o ligamentos.
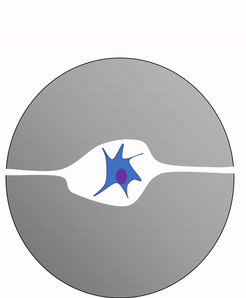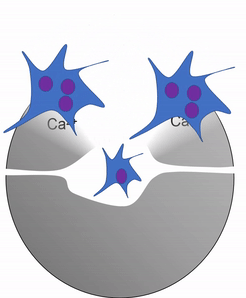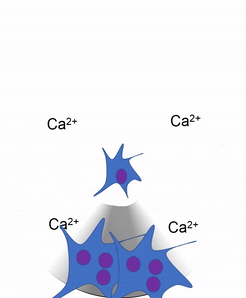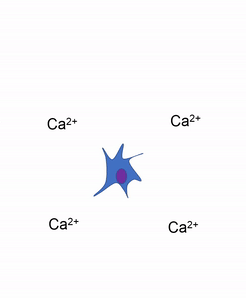
Otra célula importante que se encuentra en el periostio y en el endoste es el osteoclasto. Esta célula no se deriva de la célula osteo-condroprogenitora que da lugar a osteoblastos y osteocitos. El linaje de un osteoclasto se remonta a la médula ósea, a partir de una célula madre que también da origen a glóbulos rojos y blancos. Un osteoclasto es un pariente cercano de un glóbulo blanco llamado macrófago, más de lo que es a las células óseas. Los osteoclastos desmineralizan el tejido óseo, liberando Ca 2+ en el torrente sanguíneo. Esto es importante porque los músculos y el tejido nervioso no pueden funcionar sin Ca 2+, lo que nuestra dieta no puede proporcionar continuamente. Por lo tanto, el tejido óseo se puede considerar como un órgano de almacenamiento de calcio.

La actividad de los osteoclastos es importante durante la exfoliación de los dientes, ya que se elimina el tejido óseo para aflojar la conexión entre el hueso y el diente. La actividad de los osteoclastos también es clave para el mecanismo por el cual funciona la ortodoncia. Por último, los osteoclastos juegan un papel importante en el mantenimiento de la salud ósea, esto puede parecer contradictorio, porque los osteoclastos destruyen el tejido óseo. El tejido óseo está siendo reparado constantemente por un grupo de osteoblastos y osteoclastos que trabajan juntos, conocida como unidad de remodelación. Debido a que el tejido óseo compacto es denso, hay poco espacio para que las células funcionen. Los osteocitos pueden reparar pequeñas cantidades de daño, pero mayores cantidades de daño se acumularían con el tiempo. Para evitar esto, las unidades de remodelación trabajan constantemente para eliminar el tejido óseo y reemplazarlo con tejido óseo fresco. Las unidades de remodelación no pueden encontrar tejido óseo dañado, simplemente siguen quitando y reemplazando tejido óseo. El estrés físico en el tejido óseo hace que los osteoblastos trabajen un poco más duro, lo que conduce a un aumento de la densidad ósea. La pérdida de hormonas estimulantes de osteoblastos (como el estrógeno) puede hacer que los osteoblastos funcionen más lentamente que los osteoclastos, lo que puede disminuir la densidad ósea y potencialmente conducir a osteoporosis.

Los huesos se forman por osificación endocondral o por osificación intramembranosa. Los dos procesos son similares, la principal diferencia es que la osificación endocondral comienza con un modelo de cartílago hialino, mientras que la osificación intra-membranosa comienza con un modelo de tejido conectivo denso. La mayor parte del cráneo se desarrolla por osificación intramembranosa, excepto la mandíbula y el maxilar superior, que utilizan ambas. Por su similitud, cubrimos solo la osificación endocondral.

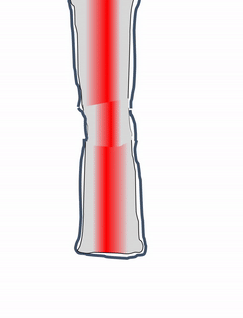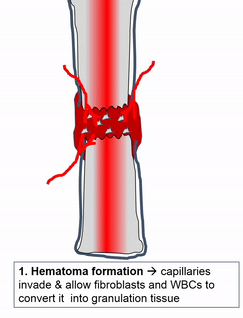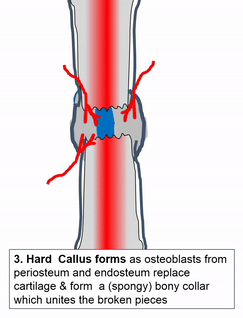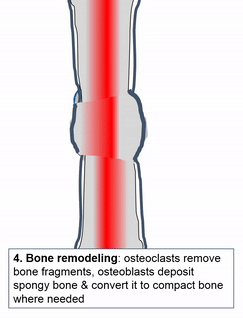
Cuando se lesiona el tejido óseo, pasa por un proceso de curación similar a la osificación endocondral. ¿Quizás recuerdas mencionar la cicatrización de heridas recapitulando el desarrollo? Aquí hay un ejemplo de un tejido adulto que reutiliza el mismo mecanismo para sanar que utilizó durante la embriogénesis para crecer en primer lugar. Similar a una lesión en la piel, el tejido óseo primero pasa por un proceso inflamatorio, y los vasos sanguíneos dañados forman coágulos. Los coágulos de sangre externos se conocen como costras, pero dentro del cuerpo se les llama hematoma. Un coágulo de sangre contiene una fibra de ECM llamada fibrina que actúa como andamio. Este andamio permite que los vasos sanguíneos crezcan en la zona, por angiogénesis. Esto a su vez permite que las células madre mesenquimales migren al área lesionada, abandonen la sangre y migren a lo largo del andamio, y se diferencien en condrocitos. Los condrocitos reemplazan el hematoma fibroso con fibrocartílago, este paso de cartílago es realmente el único paso que no tiene un paso homólogo (un paso compartido) en la reparación de heridas en la piel. A diferencia de la reparación de la piel, el crecimiento del tejido óseo necesita primero un modelo de cartílago. Con este nuevo andamio cartilaginoso, más células madre mesenquimales migran y se diferencian en osteoblastos, y reemplazan el cartílago con tejido óseo. Posteriormente, el tejido óseo esponjoso es remodelado para que coincida aproximadamente con las capas compactas y esponjosas originales. Una diferencia se puede ver después, el primer sitio de la lesión es un poco más grueso después de repararlo, similar a la forma en que el tejido de la piel puede fortalecerse después de una lesión porque las fibras de colágeno en el tejido cicatricial tienen más reticulaciones covalentes que en la TC irregular densa.
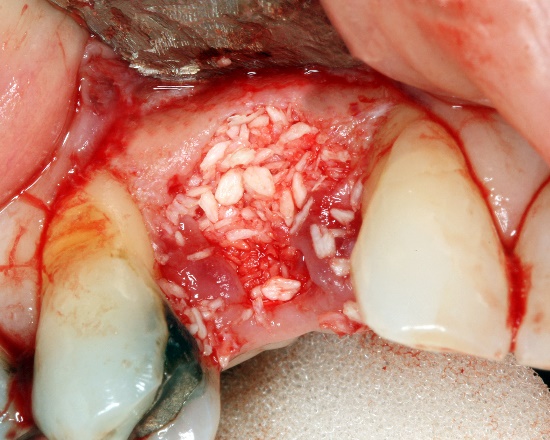
Conocer los pasos de osificación endocondral y reparación de fracturas óseas son importantes en casos en los que estimular o imitar el proceso de curación natural puede reducir la necesidad de cirugías e implantes. Por ejemplo, los pacientes pueden perder una cantidad significativa de tejido óseo con periodontitis prolongada. Si se pierde suficiente tejido óseo, un cirujano puede usar un injerto de tejido óseo para reemplazarlo. El tejido óseo de un cadáver puede ser molido, destruyendo las células pero dejando el colágeno y los minerales. Otras fuentes pueden incluir el propio tejido óseo de un paciente (un conjunto completo de costillas o fíbulas es menos importante que un conjunto intacto de huesos y dientes de la mandíbula). La MEC cosechada se extiende al sitio lesionado., donde actúa como andamio, no como reemplazo. Las células madre mesenquimales del paciente migran a través del andamio y comienzan el proceso de curación, el cual puede proceder rápidamente porque todas las materias primas están muy cerca. El andamio injertado no tiene que provenir de tejido óseo real. Puede ser dentina molida, fibrina o polímeros sintéticos impresos usando una impresora 3-D. Los andamios, sintéticos o biológicos, son reemplazados en última instancia, por lo que es posible que veas esto referido como Regeneración de Tejidos Guiada o Reabsorbible. Posteriormente aprendemos que un andamio es solo una parte del proceso de curación, hay señales importantes que pueden ser utilizadas para guiar la migración, división y diferenciación de células madre mesenquimales. Si deseas saber más, tenemos un breve video de YouTube que cubre esta nueva tecnología con más detalle, dirigido a estudiantes de Anatomía y Fisiología de 200 niveles.

Por último, es interesante considerar, desde un punto de vista evolutivo, el linaje de los cuatro tejidos mineralizados (hueso, esmalte, dentina y cemento) en humanos y otros vertebrados. ¿Uno evolucionó primero y los otros se modificaron a partir del tejido parental? La Fig. 2.31 proporciona una pista. Consideramos esta cuestión del linaje desde el punto de vista del desarrollo, en los capítulos 8-11.
![[cortar imagen de salida]](https://med.libretexts.org/@api/deki/files/22817/cut-output-image.gif)
Sangre, reticular y tejido conectivo linfático
Hay otros tejidos conectivos cubiertos en las clases de histología, aquí los cubrimos brevemente. La sangre es un tipo de tejido conectivo, compuesto por células (glóbulos rojos, en su mayoría), sustancia líquida molida (plasma sanguíneo) y fibras (fibrina). La linfa es principalmente glóbulos blancos, además de sustancia molida líquida. El tejido conectivo reticular contiene células sanguíneas, sustancia líquida molida y fibras reticulares. Se puede encontrar en órganos linfáticos donde las células sanguíneas tienden a descansar en lugar de fluir, como los ganglios linfáticos.
 Figura 2.33: Tejido cicatricial en el corazón después de un infarto de miocardio. Crédito de la imagen: “Histopatología de la cicatriz fibrosa densa reemplazando la pérdida de miocitos en el infarto de miocardio” por Katarzyna Michaud et al está licenciado bajo CC BY-SA 4.0
Figura 2.33: Tejido cicatricial en el corazón después de un infarto de miocardio. Crédito de la imagen: “Histopatología de la cicatriz fibrosa densa reemplazando la pérdida de miocitos en el infarto de miocardio” por Katarzyna Michaud et al está licenciado bajo CC BY-SA 4.0Hay otras formas de tejido conectivo que normalmente no están cubiertas en clases de histología de 200 niveles, cubrimos una de ellas: tejido cicatricial. El tejido cicatricial es hecho por fibroblastos después de una lesión. Esto ocurre después de una lesión en cualquier tejido que sea lo suficientemente grande como para que no pueda ser reemplazado rápidamente por células madre. Un ejemplo es la dentina esclerótica. El tejido cicatricial es muy similar al tejido conectivo regular denso, excepto que las fibras de colágeno en el tejido cicatricial están altamente reticuladas. Esto hace que el tejido cicatricial sea muy fuerte, pero también reduce su movilidad y capacidad para ser remodelado. El tejido cicatricial se puede formar dondequiera que haya grandes lesiones en un órgano; los fibroblastos a menudo pueden llenar el espacio más rápidamente que esperar a que ocurra la regeneración. Una vez formado, el tejido cicatricial es difícil de remodelar, por lo que es más o menos permanente.
Tejido muscular

Características generales del tejido muscular
El tejido muscular, al igual que el tejido conectivo, se deriva de somitas. La célula madre que se diferencia en uno de los diversos tipos de células musculares se llama célula mio-satélite. Dado el conjunto correcto de señales extra-celulares, las células mio-satélite se diferencian en células del músculo esquelético, liso o cardíaco. Las células del músculo esquelético son células multinucleadas largas que alguna vez fueron cientos de mioblastos que se fusionan entre sí. Al igual que un fibroblasto, un mioblasto es una célula madre que ha sufrido alguna diferenciación, suficiente para limitar su destino, y se encarga de construir el tejido adulto. En el tejido muscular, los mioblastos crean fibras proteicas que son en su mayoría intracelulares (como los filamentos delgados de actina y los filamentos gruesos de miosina). Después de fusionarse con otros mioblastos, las células musculares adultas a menudo se denominan confusamente fibras musculares, pero definitivamente son células, no moléculas como las fibras de proteínas y glicoproteínas que se encuentran dentro de la ECM del tejido conectivo. Así, también se les puede llamar miocitos. De hecho, no son la única célula multinucleada de la que lees en este capítulo, es desconcertante en cuanto a por qué esta célula suele recibir un sufijo especial, mientras que las células de osteoclastos no. En el tejido muscular adulto, las células mio-satélite permanecen, listas para someterse a mitosis y fusionarse para formar células musculares más grandes en respuesta al ejercicio o lesión. Vemos músculo esquelético en la lengua y debajo de regiones de la mucosa oral, pero no cubrimos tejido muscular liso o cardíaco.

El tejido muscular está compuesto principalmente por células musculares, conectadas por fibras de colágeno. A diferencia de las células del tejido conectivo, que pueden unirse al colágeno u otras proteínas de la ECM para la migración usando enlazadores transmembrana como las integrinas, las células musculares se anclan en su lugar a la ECM. Las células musculares expresan diferentes proteínas ligadoras para hacer esto, como la proteína Distrofina. Las capas de colágeno son delgadas entre las células, y más gruesas en la superficie externa de un músculo. Estas capas de fibras de colágeno finalmente se convierten en el denso tejido conectivo regular de un tendón. A menudo es difícil determinar dónde termina el tejido muscular y comienza el tejido conectivo, debido a las fibras de colágeno en ambos tejidos. Por extensión, también es difícil saber dónde termina el tendón y comienza el periostio. Algunos histólogos trazan líneas entre estos tejidos, pero la unión se describe mejor como una mezcla. Esto se debe a que tanto el tejido muscular como el tejido conectivo tienen el mismo linaje, del mesénquima de los somitas. Para las personas que estudian enfermedades músculo-esqueléticas como la Distrofia Muscular, existe interés en identificar las instrucciones que guían a las células madre mesenquimales para diferenciarse en células mio-satélite, especialmente considerando el mayor número de células madre mesenquimales en la médula ósea en comparación con las mio-células satélite. células satélite en el músculo de un niño o adulto. Si resultaran tratamientos novedosos de esta investigación, podría simplificar mucho los procedimientos de higiene dental para pacientes con Distrofia Muscular.
Tejido nervioso

Características generales del tejido neural
A la izquierda se muestra una imagen confocal de tejido neural. Lo que me gusta de esta imagen, y muchas veces está ausente de las imágenes de tejido neural, es que ésta muestra todas las células. La imagen típica del tejido neural utiliza manchas para resaltar las neuronas —las células especializadas en conducir electricidad— dejando fuera la glía— la mayoría de las células del sistema nervioso que hacen todo lo demás, incluyendo guiar las conexiones neuronales y cambiar la fuerza de esas conexiones cuando aprendemos. Lo que deberías ver en este momento es que el tejido neural está compuesto principalmente por células, y tiene muy poca ECM. Si bien el cerebro especialmente tiene una gran cantidad de vasos sanguíneos dentro de él, técnicamente no corren dentro del tejido neural. En cambio, hay una delgada barrera hematoencefálica que separa el cerebro de los vasos sanguíneos. Normalmente, no llamaríamos avascular al tejido neural, lo hacemos ahora solo para hacer un punto: el tejido neural comparte mucho en común con el tejido epitelial. A estas alturas, esperamos que hayas adivinado por qué: los tejidos neuronales y epiteliales comparten el mismo linaje.
3 principales tejidos embrionarios

Gastrulación
Cuando un embrión se implanta en el útero, se trata de una bola hueca de células madre idénticas y omnipotentes que se asemejan a un simple epitelio cuboidal. Pero pronto, algunas de estas células epiteliales migran hacia adentro, en un proceso muy importante llamado gastrulación. En esta etapa, tenemos dos capas distintas de células. La capa en el exterior se convierte en la epidermis, la mucosa oral y el sistema nervioso central. La capa una en el interior se convierte en el revestimiento del intestino y otros órganos huecos. A continuación, algunas celdas de la capa externa cambian. Pierden contactos con celdas vecinas, y migran hacia el medio. A medida que esto sucede, dejan de verse como células epiteliales y comienzan a parecerse a células mesenquimales. Esto produce una tercera capa de células en un embrión. Las tres capas se enumeran a continuación, junto con lo que forman en el adulto:
| 3 capas embrionarias | Destino celular |
|---|---|
| Ectodermo | Epitelio de piel y mucosa oral, tejido neural |
| Mesodermo | Tejido conectivo y muscular |
| Endodermo | Revestimiento epitelial de órganos huecos |
Cuadro 2.3: los tres tejidos embrionarios Ahora te hemos introducido furtivamente a dos términos importantes utilizados en el desarrollo. El primero es el linaje celular, lo que significa de dónde vino una célula en el pasado, y ahora el destino celular, lo que significa en lo que esa célula puede llegar a ser en el futuro. Si no pensáramos que su examen de licencia incluiría preguntas sobre los cuatro tipos principales de tejidos, nos quedaríamos con los tres tejidos embrionarios, siendo los neuronales una subfamilia de epitelios (verás por qué más adelante). Habríamos puesto tejido conectivo y muscular dentro del mismo linaje, en lugar de separarlos. Pero podemos ver por qué la academia no ha cambiado la forma en que enseña histología, la confusión que viene con el cambio no valdría la pena los beneficios que obtendríamos al agrupar las células por linaje en lugar de por apariencia. Pero esto, pensamos, es más que suficiente histología básica para un higienista dental, es hora de pasar a la histología de la cavidad oral.
Capítulo 1 * Capítulo 3