
8.2: La materia de los genes
- Page ID
- 54687

Que todas las células eucariotas contienen un núcleo se entendió a finales del siglo XIX. Para entonces, los estudios histológicos habían demostrado que los núcleos contenían en gran parte proteínas y ADN. Aproximadamente al mismo tiempo, la noción de que el núcleo contiene información genética estaba ganando terreno. En 1910, Albrecht Kossel recibió el Premio Nobel de Fisiología o Medicina 1910 por su descubrimiento de la adenina, timina, citosina y guanina (las cuatro bases de ADN), así como del uracilo en el ARN. Las leyes de herencia de Mendel, presentadas en 1865, no fueron ampliamente entendidas, probablemente porque confiaban en una fuerte dosis de aritmética y estadística, cuando no se apreciaba mucho la utilidad de la biología cuantitativa. Pero, tras el redescubrimiento tres décadas después, el número de rasgos heredados conocidos en cualquier organismo dado aumentó rápidamente. En ese momento, el ADN era conocido como una molécula pequeña y simple, compuesta solo por los cuatro nucleótidos (ver Estructura del ADN a continuación para una perspectiva histórica adicional). Entonces, la pregunta era ¿cómo podría una explicación tan pequeña y simple de la herencia de tantos rasgos físicos diferentes? El reconocimiento de que las actividades enzimáticas se heredaron de la misma manera que las características morfológicas condujo a la hipótesis de la enzima de un solo gen uno que le valió a G. W. Beadle, E. L. Tatum y J. Lederberg el Premio Nobel de Fisiología y Medicina de 1958. Cuando posteriormente se demostró que las enzimas eran proteínas, la hipótesis se convirtió en una proteína de un solo gen. Cuando se demostró que las proteínas estaban compuestas por uno o más polipéptidos, la hipótesis final se convirtió en un polipéptido de un solo gen. Sin embargo, esta relación entre genes y proteínas no arrojó ninguna luz sobre cómo el ADN podría ser el material genético. De hecho, ¡todo lo contrario! Como cadenas de hasta 20 aminoácidos diferentes, polipéptidos y proteínas tuvieron el potencial de suficiente diversidad estructural para dar cuenta del creciente número de rasgos heredables en un organismo dado. Así, las proteínas parecían más probables candidatas para las moléculas de herencia.
Los experimentos sobre los que leerás aquí comenzaron alrededor del inicio de la Primera Guerra Mundial y duraron hasta poco después de la Segunda Guerra Mundial. Durante este tiempo, aprendimos que el ADN no era un mero tetrámero, sino que de hecho era un polímero largo. Esto llevó a algunos experimentos muy inteligentes que finalmente obligaron a la comunidad científica a concluir que el ADN, no la proteína, era la molécula genética, a pesar de estar compuesta por solo cuatro unidades monoméricas. Por último, nos fijamos en el clásico trabajo de Watson, Crick, Franklin y Wilkins que reveló la estructura de la molécula genética.
A. Experimento de Griffith
Fred Neufeld, un bacteriólogo alemán que estudiaba bacterias neumocócicas a principios del siglo XX descubrió tres cepas inmunológicamente diferentes de Streptococcus pneumoniae (Tipos I, II y III). La cepa virulenta (Tipo III) fue responsable de gran parte de la mortalidad durante la pandemia de gripe española (influenza) de 1918-1920. Esta pandemia mató entre 20 y 100 millones de personas, muchas debido a que la infección viral de influenza debilitó el sistema inmunológico de los individuos infectados, haciéndolos susceptibles a la infección bacteriana por Streptococcus pneumoniae.
En la década de 1920, Frederick Griffith estaba trabajando con cepas virulentas de tipo silvestre (Tipo III) y benignas (Tipo II) de S. pneumoniae. Las dos cepas fueron fáciles de distinguir las placas de Petri debido a que la cepa virulenta creció en colonias morfológicamente lisas, mientras que la cepa benigna formó colonias ásperas. Por esta razón, las dos cepas bacterianas se denominaron S y R, respectivamente. Ahora sabemos que las células S están recubiertas con una cápsula de polisacárido (moco), haciendo que las colonias parezcan lisas. En contraste, las colonias de células R se ven ásperas (no brillan) porque carecen del recubrimiento de polisacárido.
¡Griffith sabía que inyectar células S a los ratones los mató en aproximadamente un día! Inyectando las células R no letales por otro lado, no causó ningún daño. Entonces, supuso que, tal vez, la exposición de ratones a la cepa R de S. pneumoniae primero los inmunizaría contra la infección letal por células S. Su protocolo experimental y sus resultados, publicados en 1928, se resumen a continuación.
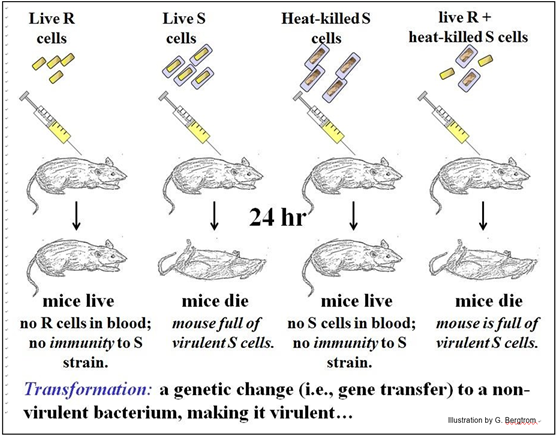
Para probar su hipótesis, Griffith inyectó a ratones células R. Algún tiempo después, les inyectó células S. Sin embargo, ¡el intento de inmunizar a los ratones contra S. pneumoniae no tuvo éxito! Los ratones control inyectados con células de la cepa S y los ratones experimentales que recibieron primero las células de la cepa R y luego las células S, ¡todos murieron en poco tiempo! Como era de esperar, los ratones inyectados con células R solo sobrevivieron.
Griffith también revisó la sangre de sus ratones para detectar la presencia de células bacterianas:
· Los ratones inyectados con células benignas de la cepa R (rugosa) sobrevivieron y después de sembrar sangre de los ratones en medio nutritivo no crecieron células bacterianas.
· Muchas colonias de células S crecieron a partir de la sangre de ratones muertos inyectados con células S.
Griffith realizó otros dos experimentos, mostrados en la ilustración:
1. Inyectó a los ratones células S muertas por calor; como era de esperar, estos ratones sobrevivieron. La sangre de estos ratones no contenía células bacterianas. Esto era “esperado” ya que calentar las células S debería tener el mismo efecto que la pasteurización tiene sobre las bacterias en la leche!
2. Griffith también inyectó a los ratones una mezcla de células R vivas y células S muertas por calor, con la esperanza de que la combinación pudiera inducir inmunidad en el ratón donde la inyección de las células R solas había fallado. Se puede imaginar su sorpresa cuando, lejos de inmunizarse, los ratones inyectados murieron y se habían acumulado abundantes células S en su sangre.
Griffith se dio cuenta de que algo importante había sucedido en sus experimentos. En la mezcla de células R vivas y células S muertas por calor, algo liberado de las células S muertas había transformado algunas células R. Griffith nombró a este “algo” el principio transformante, una molécula presente en los restos de las células S muertas y en ocasiones adquirida por unas pocas células R vivas, convirtiéndolas en células S virulentas. Ahora sabemos que las células R carecen de recubrimiento de polisacárido, y que el sistema inmunitario de la célula hospedadora puede atacar y eliminar las células R antes de que una infección grave pueda afianzarse.
B. El experimento Avery-Macleod-McCarty
Griffith no conocía la identidad química del principio transformador. Sin embargo, sus experimentos llevaron a estudios que demostraron que el ADN era “materia de genes”. Con técnicas mejoradas de purificación molecular desarrolladas en la década de 1930, O. Avery, C. MacLeod y M. McCarty transformaron células R in vitro (es decir, ¡sin la ayuda de un ratón!). Purificaron componentes de células S muertas por calor (ADN, proteínas, carbohidratos, lípidos...) y probaron por separado la capacidad transformadora de cada componente molecular en células R en un tubo de ensayo.
A continuación se resumen los experimentos de Avery et al.
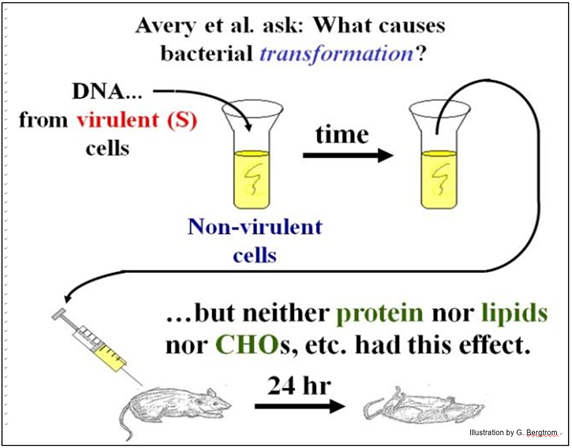
Dado que solo la fracción de ADN de las células S muertas podría causar transformación, Avery et al. concluyeron que el ADN debe ser el Principio Transformador. A pesar de estos resultados, el ADN no fue fácilmente aceptado como materia de genes. El punto de adherencia fue que el ADN estaba compuesto por sólo cuatro nucleótidos. A pesar de que los científicos sabían que el ADN era un polímero grande, todavía pensaban en el ADN como esa molécula simple, por ejemplo un polímero compuesto por secuencias repetidas de los cuatro nucleótidos:
... AGCTAGCTAGCTAGCTAGCT...
Solo las combinaciones aparentemente interminables de 20 aminoácidos en proteínas prometían la especificidad biológica necesaria para dar cuenta de los muchos rasgos genéticos de un organismo. Al carecer de diversidad estructural, el ADN se explicó como un mero andamio para genes proteicos. Para adaptar la famosa afirmación del mariscal McLuhan de que el medio es el mensaje (es decir, las ondas no se limitan a transmitir, sino que son el mensaje), muchos todavía creían que las proteínas son el medio de la información genética así como el mensaje funcional en sí mismo.
La renuencia de científicos influyentes de la época a aceptar un principio transformador del ADN privó a sus descubridores de la talla del Premio Nobel que merecía. Después de que nuevas pruebas hicieron insostenible una mayor resistencia a esa aceptación, incluso el Comité Nobel admitió que no otorgar un Premio Nobel por los descubrimientos de Avery et al. fue un error. Los experimentos clave de Alfred Hershey y Martha Chase finalmente pusieron fin a cualquier noción de que las proteínas eran genes.
167 Transformación dentro y fuera de ratones; Griffith, McCarthy et al.
C. El experimento Hershey-Chase
Bioquímicamente, se sabía que los virus bacterianos consisten en ADN encerrado en una cápsula proteica. El ciclo de vida de los virus bacterianos (bacteriófagos, o fagos para abreviar) comienza con la infección de una bacteria, como se ilustra a continuación.
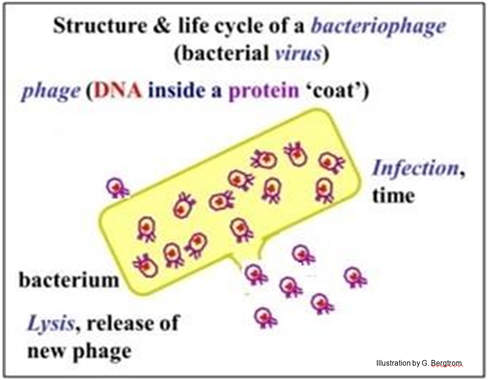
Los fagos son partículas inertes hasta que se unen e infectan a las células bacterianas. Las partículas de fago añadidas a un cultivo bacteriano se pudieron ver adheridas a superficies bacterianas en un microscopio electrónico. Los investigadores encontraron que podrían separar partículas de fagos de bacterias mediante agitación en una licuadora (similar a la que podrías tener en tu cocina). La centrifugación luego separó las células bacterianas en un sedimento en el fondo del tubo de centrífuga, dejando las partículas de fago desprendidas en el sobrenadante. Al agregar fago a las bacterias y luego separar el fago de las bacterias en diferentes momentos, fue posible determinar cuánto tiempo tuvo que permanecer adherido el fago antes de que las bacterias se infectaran. Resultó que las células sedimentadas que se habían unido al fago por tiempos cortos sobrevivirían y se reproducirían cuando se resuspendieron en medio de crecimiento. Pero las células sedimentadas que permanecían unidas al fago por tiempos más largos se habían infectado; separadas por centrifugación del fago separado y resuspendidas en medio fresco, estas células continuarían y lisaban, produciendo nuevos fagos. Por lo tanto, la transferencia de información genética para la virulencia de virus a fago tomó algún tiempo. El material genético viral responsable de la infección y virulencia aparentemente ya no se asoció con la cápsula del fago, la cual pudo ser recuperada del sobrenadante centrífugo.
Alfred Hershey y Martha Chase diseñaron un experimento para determinar si el ADN encerrado por la cápsula de proteína viral o la propia proteína de la cápsula causaron que los fagos infectaran a la bacteria. En el experimento, cultivaron por separado células de E. coli infectadas con bacteriófago T2 en presencia de 32 P o 35 S (isótopos radiactivos de fósforo y azufre, respectivamente). El resultado fue producir fagos que contenían ADN radiactivo o proteínas radiactivas, pero no ambas (recordemos que solo el ADN contiene fósforo y solo las proteínas contienen azufre). Luego infectaron por separado células frescas de E. coli con fagos radiactivos marcados con 32 P o 35 S. Su experimento se describe a continuación.
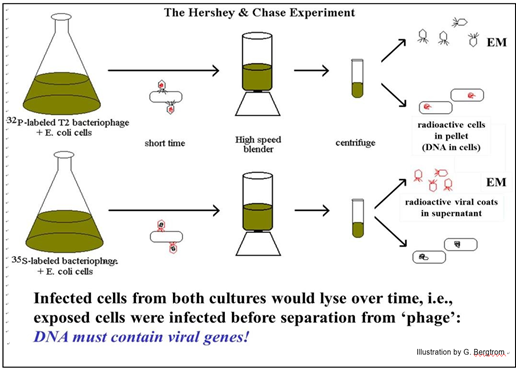
Los fagos y las células se incubaron con 32P o 35S el tiempo suficiente para permitir la infección. Parte de cada cultivo se dejó continuar y lisar para demostrar que las células estaban infectadas. El resto de cada mezcla se envió a la licuadora. Después de la centrifugación de cada mezcla, se examinaron los sedimentos y los sobrenadantes para ver a dónde habían ido las proteínas radiactivas o el ADN. De los resultados, el 32P siempre terminó en el sedimento de células bacterianas mientras que el 35S se encontró en el remanente de fago en el sobrenadante. Hershey y Chase concluyeron que el material genético de los virus bacterianos era ADN y no proteína, así como Avery et al. habían sugerido que el ADN era el principio de transformación bacteriana.
Dada la resistencia anterior a que el ADN “simple” sea el material genético, Hershey y Chase utilizaron un lenguaje cauteloso para enmarcar sus conclusiones. No es necesario tenerlos; todos los experimentos posteriores confirmaron que el ADN era el material genético. Simultáneamente con estas confirmaciones hubo experimentos que demostraban que el ADN podría no ser (de hecho, no lo era) ¡una molécula tan simple y sin complicaciones! Por sus contribuciones finales para fijar el ADN como “materia de genes”, Alfred D. Hershey compartió el Premio Nobel de Fisiología o Medicina 1969 con Max Delbruck y Salvador E. Luria.
168 Hershey y Chase: los genes virales están en el ADN viral