
9.2: Replicación de ADN
- Page ID
- 54517

Como hemos visto, las cadenas de ADN tienen direccionalidad, con un 5' nucleótido-fosfato y un extremo 3' desoxirribosa hidroxilo. Esto es incluso cierto para los cromosomas bacterianos circulares..., ¡si se rompe el círculo! Debido a que las hebras de la doble hélice son antiparalelas, el extremo 5' de una hebra se alinea con el extremo 3' de la otra en ambos extremos de la doble hélice. El emparejamiento complementario de bases en el ADN significa que la secuencia de bases de una cadena se puede utilizar como molde para hacer una nueva cadena complementaria. Como veremos, esta estructura del ADN creó algunos dilemas interesantes para entender la bioquímica de la replicación. El desconcierto que rodea cómo procede la replicación comienza con experimentos que visualizan la replicación del ADN.
A. Visualización de bifurcación de replicación y replicación
Recordemos que el fenómeno de conjugación bacteriana permitió una demostración de que los cromosomas bacterianos eran circulares. En 1963, John Cairns confirmó este hecho mediante la visualización directa del ADN bacteriano. Cultivó células de E. coli durante largos periodos en 3Htimidina (3H-T) para hacer radioactivo todo su ADN celular. Luego interrumpió las células suavemente para minimizar el daño al ADN. Se permitió que el ADN liberado se asentara y se adhiriera a las membranas. Se colocó una película sensible sobre la membrana y se dejó tiempo para que la radiación expusiera la película. Después de que Cairns desarrolló las autorradiografías, examinó los resultados en el microscopio electrónico. Vio huellas de granos de plata en las autorradiografías (el mismo tipo de granos de plata que crean una imagen en película en la fotografía anticuada). Mira los dos dibujos de sus autorradiografías en la página siguiente.
Cairns midió la longitud de las pistas “plateadas”, que generalmente consistían en tres posibles bucles cerrados, o círculos. Las circunferencias de dos de estos círculos fueron siempre iguales, su longitud predicha de cerca por el contenido de ADN de una sola célula no divisoria. Por lo tanto, Cairns interpretó estas imágenes como ADN bacteriano en el proceso de replicación. A continuación se ilustran las autorradiografías de Cairns y las medidas que lo llevaron a concluir que estaba mirando imágenes de cromosomas circulares bacterianos.

Arregló sus imágenes de autorradiografía en una secuencia (abajo) para hacer su punto.
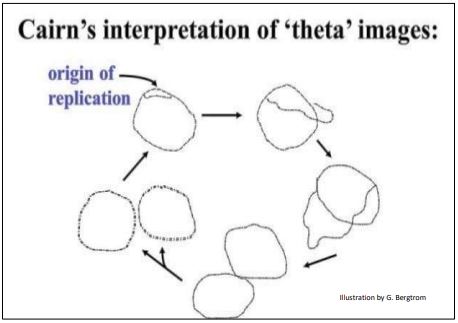
Porque los cromosomas replicantes se veían (¡vagamente!) como la letra griega \(\theta\), Cairns los llamó theta images. Inferyó que la replicación comienza en un único origen de replicación en el cromosoma bacteriano, procediendo alrededor del círculo hasta su finalización.
Experimentos posteriores de David Prescott demostraron replicación bidireccional..., esa replicación efectivamente comenzó en un origen de replicación, después de lo cual la doble hélice se desenrolló y replicó en ambas direcciones, lejos de los orígenes, formando dos horquillas de replicación (ilustrado abajo).
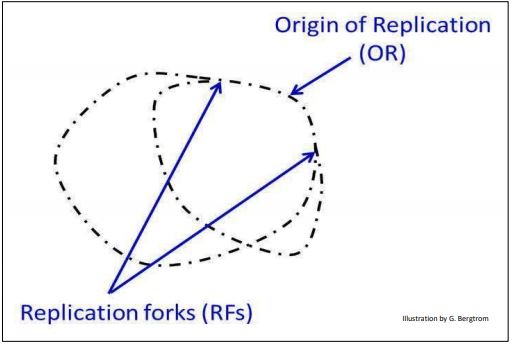
176 Replicación bidreccional semiconservativa a partir de dos RFs
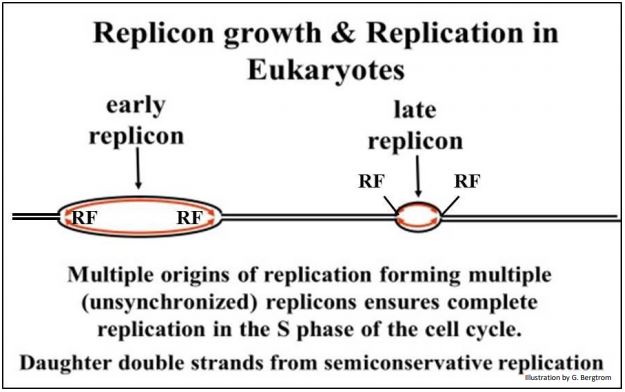
Las células bacterianas pueden dividirse cada hora (o incluso menos); la tasa de síntesis de ADN bacteriano es de aproximadamente 2 X 106 pares de bases por hora. Un núcleo celular eucariota típico contiene miles de veces más ADN que una bacteria, y las células eucariotas típicas se duplican cada 15-20 horas. Incluso un cromosoma pequeño puede contener cientos o miles de veces más ADN que una bacteria. ¡Parecía que las células eucariotas no podían darse el lujo de duplicar su ADN a una velocidad de replicación bacteriana! Los eucariotas resolvieron este problema no evolucionando una bioquímica de replicación más rápida, sino utilizando múltiples orígenes de replicación a partir de los cuales procede la síntesis de ADN en ambas direcciones. Esto da como resultado la creación de múltiples replicones.
Cada replicón se agranda y finalmente se encuentra con otros replicones en crecimiento en cada lado para replicar la mayor parte de cada cromosoma lineal, sugerido en la siguiente ilustración.
Antes de considerar en detalle los eventos bioquímicos en las horquillas de replicación, veamos el papel de las enzimas ADN polimerasa en el proceso.
B. Las ADN polimerasas catalizan la replicación
La primera de estas enzimas fue descubierta en E. coli por Arthur Kornberg, por lo que recibió el Premio Nobel de Química de 1959. ¡Thomas Kornberg, uno de los hijos de Arthur más tarde encontró dos ADN polimerasas más! Todas las ADN polimerasas requieren una cadena molde contra la cual sintetizar una nueva cadena complementaria. Todos ellos cultivan nuevo ADN añadiendo al extremo 3' de la cadena de ADN en crecimiento en sucesivas reacciones de condensación. Y finalmente, todas las ADN polimerasas también tienen la extraña propiedad que solo pueden agregar a una cadena preexistente de ácido nucleico, ¡planteando la cuestión de dónde viene la hebra 'preexistente'! Las ADN polimerasas catalizan la formación de un enlace fosfodiéster entre el extremo de una cadena en crecimiento y el nucleótido entrante complementario a la cadena molde. La energía para la formación del enlace fosfodiéster proviene en parte de la hidrólisis de dos fosfatos (pirofosfato) del nucleótido entrante durante la reacción. Si bien la replicación requiere la participación de muchas proteínas nucleares tanto en procariotas como eucariotas, las ADN polimerasas realizan los pasos básicos de replicación, como se muestra en la siguiente ilustración.
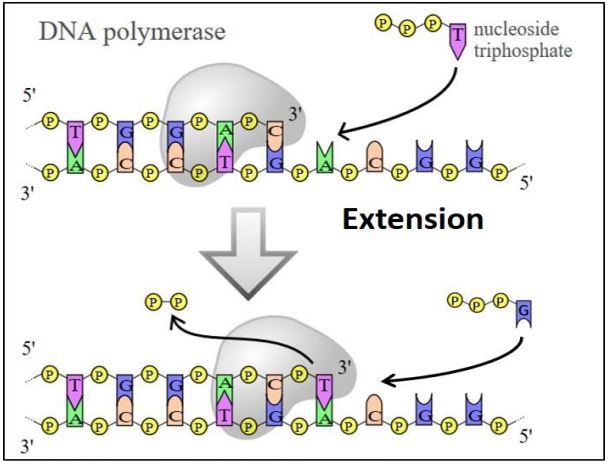
178 ADN polimerasas y sus actividades
Aunque las ADN polimerasas replican ADN con alta fidelidad con tan solo un error por 107 nucleótidos, ocurren errores. La capacidad de corrección de algunas ADN polimerasas corrige muchos de estos errores. La polimerasa puede detectar un par de bases desapareadas, ralentizar y luego catalizar hidrolizaciones repetidas de nucleótidos hasta que alcanza el par de bases desapareadas. Esta corrección básica por ADN polimerasa se muestra a continuación.
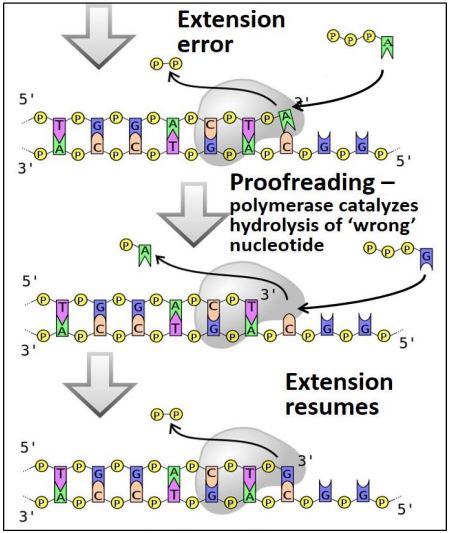
Después de reparar el desajuste, la ADN polimerasa reanuda el movimiento hacia adelante Por supuesto, no todos los errores son captados por este u otros mecanismos de reparación (ver Reparación de ADN, a continuación). Las mutaciones en las células de la línea germinal eucariota que eluden la corrección pueden causar enfermedades genéticas. Sin embargo, la mayoría son las mutaciones que alimentan la evolución. Sin mutaciones en las células de la línea germinal (óvulo y esperma), no habría mutaciones ni evolución, y sin evolución, ¡la vida misma habría llegado a un rápido callejón sin salida! Otros errores de replicación pueden generar mutaciones en células somáticas. Si estas mutaciones somáticas escapan a la corrección, pueden tener graves consecuencias, entre ellas la generación de tumores y cánceres.
C. El Proceso de Replicación
La replicación del ADN es una secuencia de reacciones repetidas de condensación (síntesis de deshidratación) que unen monómeros de nucleótidos en un polímero de ADN. Como todas las polimerizaciones biológicas, la replicación procede en tres etapas enzimáticamente catalizadas y coordinadas: iniciación, elongación y terminación.
1. Iniciación
Como hemos visto, la síntesis de ADN comienza en uno o más orígenes o replicación. Estas son secuencias de ADN dirigidas por proteínas iniciadoras en E. coli (abajo).
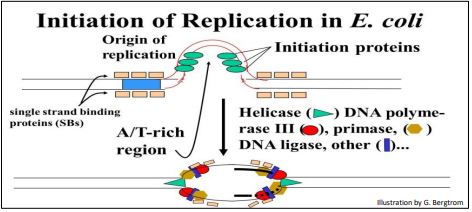
Después de romper los enlaces de hidrógeno en el origen de la replicación, la doble hélice del ADN es progresivamente descomprimida en ambas direcciones (es decir, por replicación bidireccional). Las hebras de ADN separadas sirven como plantillas para la nueva síntesis de ADN. Las secuencias en los orígenes de replicación que se unen a proteínas de iniciación tienden a ser ricas en bases de adenina y timina. Esto se debe a que los pares de bases A-T tienen dos enlaces de hidrógeno (H-) que requieren menos energía para romperse que los tres enlaces H que mantienen los pares G-C juntos. Una vez que las proteínas de iniciación aflojan los enlaces H en un origen de replicación, la ADN helicasa utiliza la energía de la hidrólisis de ATP para desenrollar la doble hélice. La ADN polimerasa III es la principal enzima que luego alarga el nuevo ADN. Una vez iniciada, se forma una burbuja de replicación (replicón) a medida que transcurren ciclos repetidos de elongación en horquillas de replicación opuestas
179 Iniciación de replicación en E. coli
Recordando que solo se pueden agregar nuevos nucleótidos al grupo hidroxilo 3' libre de una cadena de ácido nucleico preexistente. Dado que ninguna ADN polimerasa conocida puede comenzar a sintetizar nuevas cadenas de ADN desde cero, ¡esto es un problema! Por lo tanto, la acción de las ADN polimerasas requiere de un cebador, una cadena de ácido nucleico a la que añadir nucleótidos. Las preguntas fueron..., ¿cuál es la cartilla y de dónde viene? Dado que las ARN polimerasas (enzimas que catalizan la síntesis de ARN) son la única nucleotídica polimerasa que puede hacer crecer una nueva cadena de ácido nucleico contra un molde de ADN desde cero (es decir, desde la primera base), se sugirió que el ARN podría ser el cebador, Después de la síntesis de un cebador de ARN corto, nuevo los desoxinucléotidos serían añadidos a su extremo 3' por la ADN polimerasa. El descubrimiento de tramos cortos de nucleótidos de ARN en el extremo 5' de los fragmentos de Okazaki confirmó la noción de cebadores de ARN. Ahora sabemos que las células utilizan primasa, una ARN polimerasa especial activa durante la replicación, para hacer esos cebadores de ARN contra plantillas de ADN antes de que una ADN polimerasa pueda hacer crecer las cadenas de ADN en las horquillas de replicación. Como veremos ahora, el requisito de cebadores de ARN no está en ninguna parte más en evidencia en eventos en una bifurcación de replicación.
2. Alargamiento
Al observar el alargamiento en una horquilla de replicación (abajo), vemos otro problema:
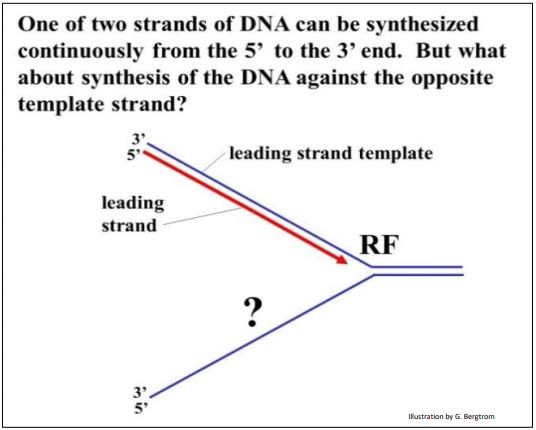
Una de las dos nuevas cadenas de ADN puede crecer continuamente hacia la horquilla de replicación a medida que la doble hélice se desenrolla. Pero, ¿qué pasa con la otra hebra? O esta otra hebra debe crecer en pedazos en sentido contrario, o debe esperar para comenzar la síntesis hasta que la doble hélice esté completamente desenrollada. Si una cadena de ADN debe replicarse en fragmentos, entonces esos fragmentos tendrían que ser cosidos (es decir, ligados) juntos. El problema se ilustra a continuación.
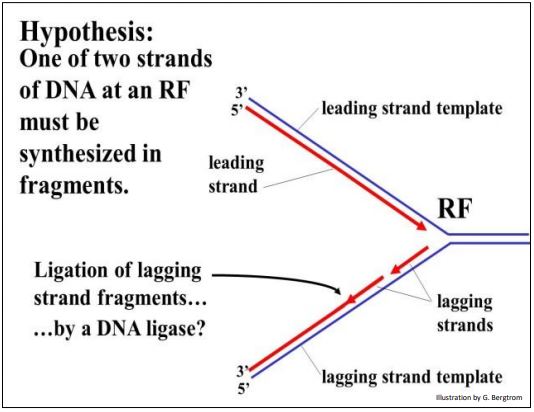
Según esta hipótesis, una nueva cadena líder de ADN se alarga continuamente mediante la adición secuencial de nucleótidos a su extremo 3' contra su molde de cadena principal. La otra cadena, sin embargo, estaría hecha en piezas que se unirían en enlaces fosfodiéster en una reacción posterior. Debido al paso extra y presumiblemente el tiempo extra que se necesita para hacer y unir estos nuevos fragmentos de ADN, este nuevo ADN se llama la hebra rezagada, haciendo de su plantilla la plantilla de hebra rezagada.
Reiji Okazaki y sus colegas estaban estudiando mutantes del fago T4 que crecieron lentamente en sus células hospedadoras de E. coli. Graficaron las tasas de crecimiento del fago T4 de tipo silvestre y mutante y demostraron que el crecimiento lento se debió a una enzima ADN ligasa deficiente, ya conocida por catalizar la circularización de moléculas de ADN de fagos lineales replicadas en células hospedadoras infectadas. La gráfica a continuación resume sus resultados.
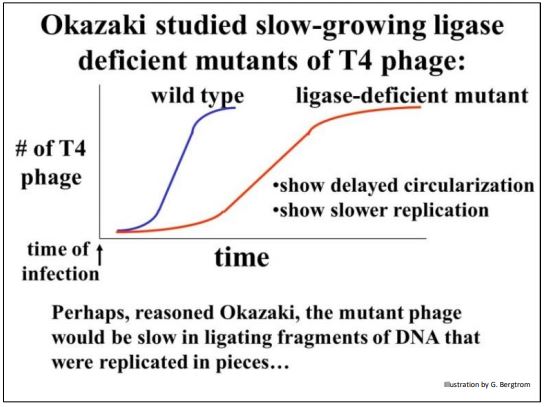
La hipótesis de Okazaki fue que la ADN ligasa deficiente en el fago mutante no solo ralentizó la circularización del ADN del fago T4 replicante, sino que también sería lenta en la unión de fragmentos de ADN de fago replicados contra al menos una de las dos cadenas de ADN molde. Cuando se probó la hipótesis, los Okazakis encontraron que fragmentos cortos de ADN efectivamente se acumulaban en células de E. coli infectadas con mutantes deficientes en ligasa, pero no en células infectadas con fagos de tipo silvestre. Los fragmentos de hebra rezagados ahora se llaman fragmentos de Okazaki.
180 Experimentos y fragmentos de Okazaki - Resolviendo un problema en una RF
Se elaboran 181 fragmentos de Okazaki comenzando con cebadores de ARN
Puedes consultar la investigación original de Okazaki en este enlace.
Cada fragmento de Okazaki tendría que comenzar con un cebador de ARN 5', ¡creando otro dilema más! El cebador de ARN debe reemplazarse con desoxinucleótidos antes de unir los fragmentos. Esto de hecho sucede, y el proceso que se ilustra a continuación
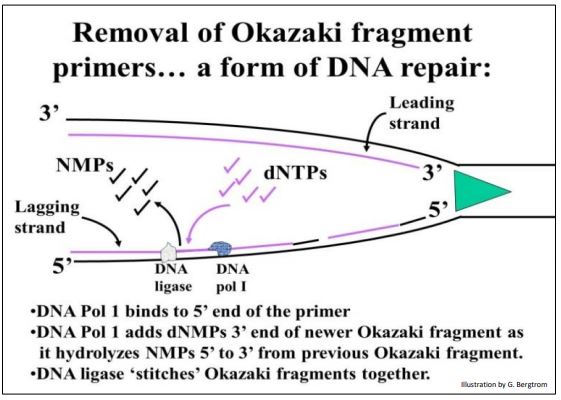
La eliminación de nucleótidos cebadores de ARN de fragmentos de Okazali requiere la acción de la ADN polimerasa I, una enzima que también puede catalizar la hidrólisis de los enlaces fosfodiéster entre los nucleótidos de ARN (o ADN) del extremo 5' de una cadena de ácido nucleico. La endonucleasa colgajo 1 (FEN 1) también juega un papel en la eliminación de “colgajos” de ácido nucleico de los extremos 5' de los fragmentos a menudo desplazados por la polimerasa ya que reemplaza al cebador de replicación. Al mismo tiempo que se eliminan los nucleótidos de ARN, la ADN polimerasa I cataliza su reemplazo por los desoxinucleótidos apropiados. Finalmente, cuando un fragmento es completamente ADN, la ADN ligasa lo une al resto del ADN de la hebra retrasada ya ensamblada. Debido a su actividad exonucleasa 5' (no encontrada en otras ADN polimerasas), la ADN polimerasa 1 también juega un papel único en la reparación del ADN (discutido más adelante). Como sugirió Cairn y otros demostraron, la replicación procede en dos direcciones desde el origen para formar un replicón con sus dos bifurcación de replicación (RFs). Cada RF tiene una primasa asociada con la replicación de fragmentos de Okazaki a lo largo de plantillas de hebras rezagadas.
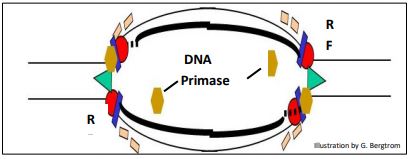
Como sugirió Cairn y otros demostraron, la replicación procede en dos direcciones desde el origen para formar un replicón con sus dos bifurcación de replicación (RFs). Cada RF tiene una primasa asociada con la replicación de fragmentos de Okazaki a lo largo de plantillas de hebras rezagadas.
A continuación se muestra el requisito de primasas en las horquillas de replicación.
Ahora podemos preguntarnos qué pasa cuando los replicones llegan a los extremos de los cromosomas lineales en eucariotas.
3. Terminación
En procariotas, la replicación se completa cuando dos horquillas de replicación se encuentran después de replicar su porción de la molécula circular de ADN. En eucariotas, muchos replicones se fusionan para convertirse en replicones más grandes, llegando finalmente a los extremos de los cromosomas..., donde hay otro problema más (abajo)!
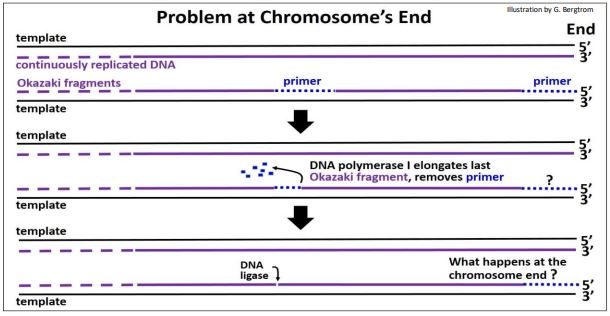
Cuando un replicón se acerca al final de un cromosoma (es decir, una molécula de ADN bicatenario), la cadena sintetizada se detiene continuamente cuando alcanza el extremo 5' de su ADN molde. En teoría, la síntesis de un último fragmento de Okazaki puede cebarse desde el extremo 3' de la cadena molde rezagada. La ilustración anterior implica la eliminación de un cebador del penúltimo fragmento de Okazaki y el reemplazo catalizado por ADN polimerasa con nucleótidos de ADN. Pero ¿qué pasa con el último fragmento de Okazaki? ¿Se hidrolizaría su imprimación? Además, sin un extremo 3' libre para agregar, ¿cómo se reemplazan esos nucleótidos de ARN con nucleótidos de ADN? El problema aquí es que cada vez que una célula se replica, una hebra de nuevo ADN (probablemente ambas) se acortaría cada vez más. Por supuesto, esto no serviría..., ¡y no sucede! La replicación eucariota se somete a un proceso de terminación que implica extender la longitud de una de las dos cadenas por la enzima telomerasa. La acción de la telomerasa se resume en la siguiente ilustración
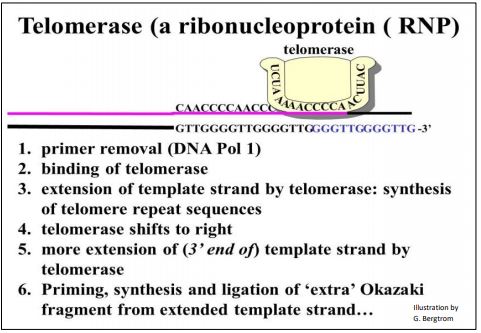
La telomerasa consiste en varias proteínas y un ARN. A partir del dibujo,, el componente de ARN sirve como molde para la extensión 5'-> 3' de la cadena de ADN problemática. La proteína con la actividad transcriptasa inversa requerida se llama Te lomerasa R everse T ranscriptasa, o TERT. El omponente Te lomerasa R NA C se llama TERC. Carol Greider, Jack Szostak y Elizabeth Blackburn compartieron el Premio Nobel de Fisiología o Medicina 2009 por descubrir la telomerasa.
183 La replicación de la telomerasa previene el acortamiento cromosómico https: //youtu.be/m4dmfrxgkku
Una de las observaciones recientes más interesantes fue que las células diferenciadas y no divisorias ya no producen la enzima telomerasa. Por otro lado, los genes de la telomerasa son activos en células normales en división (por ejemplo, células madre) y células cancerosas, las cuales contienen abundante telomerasa.
4. ¿La replicación es processiva?
Los dibujos de replicones y horquillas de replicación sugieren eventos separados en cada cadena de ADN. Sin embargo, los eventos en bifurcación de replicación parecen estar coordinados. La replicación puede ser processiva, lo que significa que ambas nuevas cadenas de ADN se replican en la misma dirección al mismo tiempo, suavizando el proceso. ¿Cómo podría ser esto posible? El dibujo a continuación muestra la flexión del ADN de la plantilla de hebra rezagada, de manera que mira en la misma dirección que la cadena principal en la horquilla de replicación.
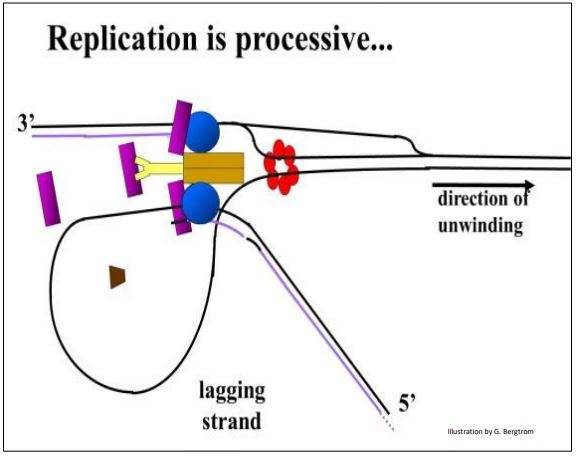
La estructura del replisoma cartografiada en la horquilla de replicación consiste en proteínas clamp, primasa, helicasa, ADN polimerasa y proteínas de unión monocatenarias entre otras.
Las técnicas más nuevas de visualización de la replicación mediante videografía de fluorescencia en tiempo real cuestionan el modelo processivo, lo que sugiere que el proceso de replicación es todo menos fluido. ¿De hecho no están coordinadas la replicación de las cadenas rezagadas y líderes? Alternativamente, ¿es el movimiento entrecortado del alargamiento del ADN en el video un artefacto, de manera que el modelo de replicación suave y coordinada integrado en un replisoma sigue siendo válido? ¿O la coordinación se define y se logra de alguna otra manera? Consulta el video tú mismo en el artículo aquí.
5. Un problema más con la replicación
Cairns grabó muchas imágenes de E.coli del tipo que se muestra a continuación.

La apariencia enrollada y retorcida de los círculos replicantes se interpretó como una consecuencia natural de tratar de separar hebras de ADN entrelazadas helicoidalmente... ¡o hebras entrelazadas de cualquier material! A medida que las hebras continuaban desenrollándose, el ADN debería torcerse en una superbobina de ADN. El aumento del desenrollamiento del ADN provocaría que los enlaces fosfodiéster en el ADN se rompan, fragmentando el ADN. Obviamente, esto no sucede. Se diseñaron experimentos para demostrar superenrollamiento y para probar hipótesis explicando cómo las células relajan las superbobinas durante la replicación. Las pruebas de estas hipótesis revelaron las enzimas topoisomerasas. Estas enzimas se unen y se mantienen al ADN, catalizan la hidrólisis de enlaces fosfodiéster, controlan el desenrollado de la doble hélice y finalmente catalizan la reformación de los enlaces fosfodiéster. Es importante señalar que las topoisomerasas no forman parte de un replisoma, sino que pueden actuar lejos de una bifurcación de replicación, probablemente respondiendo a las tensiones en el ADN sobreenrollado. Recordemos que las topoisomerasas comprenden gran parte de la proteína que se encuentra junto a la cromatina eucariota.
185 topoisomerasas alivian el superenrollamiento durante la replicación
Hemos considerado a la mayoría de los actores moleculares en replicación. A continuación se muestra una lista de las proteínas de replicación clave y sus funciones (a partir de aquí).
| Enzima | Función en la replicación del ADN |
| Helicasa de ADN | También conocida como enzima desestabilizadora de hélice. Desenrolla la doble hélice de ADN en el Tenedor de Replicación. |
| ADN Polimerasa |
Construye una nueva cadena de ADN dúplex mediante la adición de nucleótidos en la dirección 5' a 3'. También realiza corrección de pruebas y corrección de errores. |
| Pinza de ADN | Proteína que impide que la ADN polimerasa III se disocie de la cadena parental de ADN. |
| Proteínas de unión monocatenaria (SSB) | Se une al ADNss y evita que la doble hélice del ADN vuelva a aparearse después de que la helicasa de ADN la desenrolle, manteniendo así la separación de cadenas. |
| Topoisomerasa | Relaja el ADN de su naturaleza súper enrollada. |
| ADN girasa | Alivia la cepa de desenrollamiento por ADN helicasa; este es un tipo específico de topoisomerasa |
| ADN Ligasa | Vuelva a recocer las hebras semiconservadoras y se une a los fragmentos de Okazaki de la hebra rezagada |
| Primase | Proporciona un punto de partida de ARN (o ADN) para que la ADN polimerasa comience la síntesis de la nueva cadena de ADN. |
| Telomerasa | Alarga el ADN telomérico añadiendo secuencias repetitivas de nucleótidos a los extremos de los cromosomas eucariotas. |