
9.3: Reparación de ADN
- Page ID
- 54533

A. El daño en el ADN (mutación) es un hecho de la vida
Generalmente aceptamos la noción de que la replicación duplica fielmente el material genético. Al mismo tiempo, la evolución no sería posible sin mutación, y la mutación no es posible sin al menos algunas consecuencias adversas.
Las mutaciones de la línea germinal son heredables. Cuando están presentes en uno, pero especialmente en ambos alelos de un gen, tales mutaciones pueden resultar en enfermedades genéticas (por ejemplo, enfermedad de Tay-Sach, fibrosis quística, hemofilia, anemia falciforme, etc.). En lugar de causar enfermedades, algunas mutaciones de la línea germinal pueden aumentar la probabilidad de enfermarse (por ejemplo, las mutaciones del gen BRCA2 aumentan en gran medida las probabilidades de que una mujer contraiga cáncer de mama). Las mutaciones somáticas en células que se dividen activamente pueden dar como resultado “quistes” benignos o tumores malignos (es decir, cáncer). Otras mutaciones somáticas pueden desempeñar un papel en la demencia (enfermedad de Alzheimer) o en algunas de las neuropatologías a lo largo del espectro autista.
Dado que la compleja química de replicación está sujeta a una alta tasa de error inherente, las células han desarrollado sistemas de reparación de ADN para sobrevivir a altas tasas de mutación. Como vimos, las propias ADN polimerasas tienen capacidad correctora para que las bases insertadas incorrectamente puedan eliminarse y reemplazarse rápidamente. Más allá de esto, múltiples mecanismos han evolucionado para reparar pares de bases no coincidentes y otros tipos de ADN dañado que escapan a la detección temprana. La frecuencia y dónde ocurre el daño en el ADN es aleatorio, como es qué daño se reparará y cuál se escapará para convertirse en una mutación. Para quienes sufren las terribles consecuencias de una mutación no reparada, el equilibrio entre ADN retenido y reparado, daño es por decir lo menos, imperfecto. Sin embargo, la evolución y la continuidad de la vida misma se basan en este equilibrio.
B. Qué Causa el Daño al ADN
El ADN está más expuesto y por lo tanto más vulnerable al daño, especialmente en eucariotas. El daño más simple al ADN durante la replicación es la mutación puntual, la inserción accidental de un nucleótido 'incorrecto' en una cadena de ADN en crecimiento. Otras mutaciones, igualmente accidentales, incluyen deleciones de ADN, duplicaciones, inversiones, etc., cualquiera de las cuales podría escapar a la reparación. Las causas del daño al ADN pueden ser químicas o físicas, e incluyen eventos intracelulares espontáneos (por ejemplo, reacciones oxidativas) y factores ambientales (radiación, químicos exógenos, etc.). Con base en estudios de diferentes tipos de daño en el ADN, Thomas Lindahl estimó que los eventos que dañan el ADN podrían estar ocurriendo a razón de ¡10,000 por día! Lindahl se dio cuenta de que debe haber algunos “mecanismos fundamentales de reparación del ADN” en el trabajo para proteger a las células contra una tasa tan alta de daño en el ADN. El descubrimiento del mecanismo de reparación de escisión de base le valió a Thomas Lindahl una participación en el Premio Nobel de Química 2015. Los factores ambientales que pueden dañar el ADN incluyen la luz UV, los rayos X y otras radiaciones, así como los químicos (por ejemplo, toxinas, carcinógenos e incluso drogas, etc.). Tanto las células germinales como las somáticas pueden verse afectadas. Si bien las mutaciones pueden causar y causan enfermedades a menudo debilitantes, es instructivo mantener el impacto de las mutaciones en perspectiva. La mayoría de las mutaciones son en realidad silenciosas; no causan enfermedad. Además, se repara mucho daño en el ADN. Las células corrigen más del 99.9% de los cambios de base equivocados antes de que tengan la posibilidad de convertirse en mutaciones. Por eso pensamos en la replicación como un proceso “fiel”. Veamos algunos tipos comunes de daño en el ADN que generalmente se reparan:
- Dímeros de pirimidina, típicos de las timinas adyacentes (con menos frecuencia citosinas) en una sola cadena de ADN, causados por la exposición UV
- Depuración; la eliminación hidrolítica de guanina o adenina del #1 C (carbono) de desoxirribosa en una cadena de ADN
- Desaminación: eliminación hidrolítica de grupos amino (-NH2) de guanina (más común), citosina o adenina
- Daño oxidativo de la desoxirribosa con cualquier base, pero más comúnmente purinas
- Metilación inapropiada de cualquier base, pero más comúnmente purinas
- Rotura de la cadena de ADN durante la replicación o por radiación o exposición química
C. Algunas consecuencias moleculares del daño no corregido en el ADN
Si bien las bacterias sufren daños en el ADN, aquí nos centraremos en los eucariotas ya que han desarrollado los mecanismos más sofisticados. Recuerde que el daño en el ADN no reparado se transmitirá a las células hijas en la mitosis, o podría transmitirse a la siguiente generación si la mutación ocurre en una célula germinal.
A continuación, consideremos algunas consecuencias moleculares del daño del ADN no corregido.
1. Depuración
Esta es la eliminación hidrolítica espontánea de guanina o adenina de la desoxirribosa C #1 en una cadena de ADN. ¡Su frecuencia de 5000 depuraciones por célula al día enfatiza la alta tasa de daño en el ADN que exige una solución! Si no se repara, la depuración da como resultado una deleción de un solo par de bases en un cromosoma después de la replicación, dejando el ADN en la misma región del otro cromosoma sin cambios. A continuación se ilustran los efectos de la depuración.
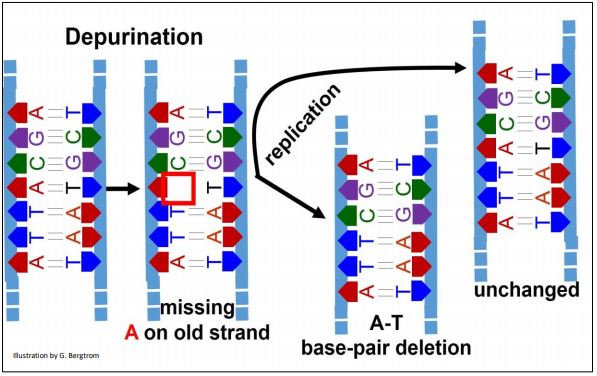
El replisoma ignora la base faltante durante la replicación de la región de ADN depurado (una A en este ejemplo), saltando a la C en el ADN molde depurado. Sin reparar, un nuevo ADN bicatenario tendrá una deleción, dejando al otro nuevo sin mutación.
2. Dimerización de pirimidina
La exposición a la luz UV del ADN puede causar que las pirimidinas adyacentes (comúnmente timinas; con menos frecuencia, citosinas) en una cadena de ADN se dimericen. ¡Los dímeros de pirimidina se forman a una velocidad de un poco menos de 100 por célula al día!
La dimerización no corregida da como resultado una deleción de 2 bases en un cromosoma mientras que el otro no cambia (abajo).
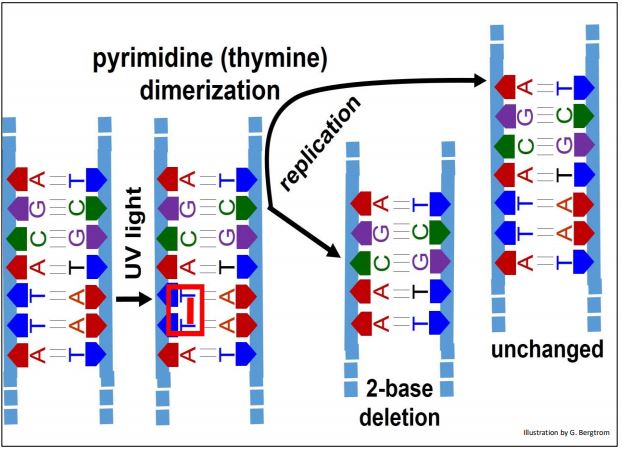
Se puede predecir que la corrección de este daño inducido por radiación implicará la alteración de los dímeros (en este caso dímeros de timina), o la eliminación y reemplazo de las bases dimerizadas por bases monoméricas.
3. Desaminación
La desaminación es la eliminación hidrolítica de grupos amino (-NH2) de guanina (más común), citosina o adenina, a una tasa de 100 por célula por día. La desaminación no afecta a la timina (¡porque no tiene grupos —amino!). La desaminación no corregida da como resultado una sustitución de bases en un cromosoma (en realidad, una sustitución de pares T-A por el C-G original en este ejemplo) y ningún cambio en el otro. La desaminación de adenina o guanina da como resultado bases no naturales (hipoxantina y xantina, respectivamente). Estos son fácilmente reconocidos y corregidos por los sistemas de reparación de ADN. El par de bases U-A permanece ocasionalmente sin reparar. A continuación se muestran las consecuencias de la desaminación a la secuencia de bases.
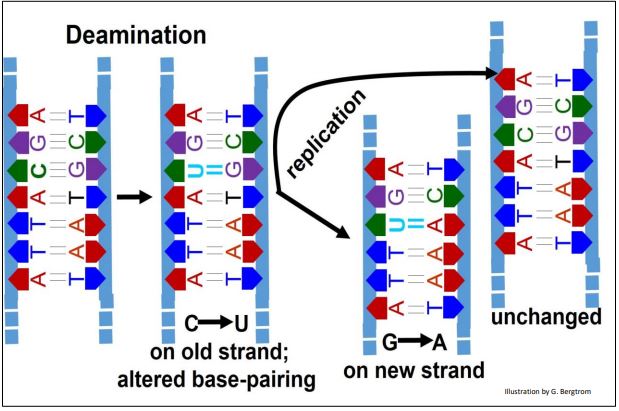
D. Mecanismos de Reparación del ADN
Muchas enzimas y proteínas están involucradas en la reparación del ADN. Algunos de estos funcionan en replicación normal, mitosis y meiosis, pero fueron cooptados por actividades de reparación del ADN. Estas cooptaciones moleculares son tan vitales para la función celular normal que algunas actividades de reparación y actores moleculares están altamente conservados en la evolución. Entre las diferentes vías de reparación del ADN que se han identificado, analizaremos la Reparación de Escisión de Base, la Reparación de Escisión de Nucleótidos, la Reparación Acoplada a la Transcripción, la Unión de Extremos No Homóloga y la Recombinación Homóloga (de estas, la quizás el más complejo).
1. Reparación de Escisión Base
Al detectar y reconocer una base incorrecta (por ejemplo, bases oxidadas, bases de apertura, Cs o As desaminadas, bases que contienen enlaces C=C saturados a enlaces C-C...), las ADN glicosilasas específicas catalizan la hidrólisis de la base dañada de la desoxirribosa afectada en el ADN. Para conocer más sobre las versiones específicas de esta enzima, haga clic aquí. A continuación se resumen los eventos de reparación por escisión de bases.
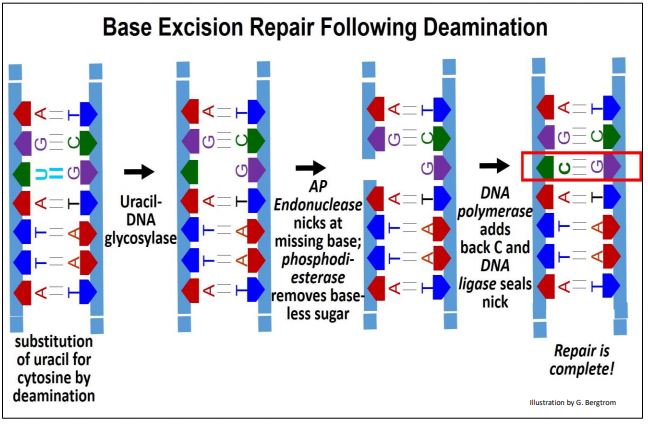
Después de que una ADN glicosilasa elimina una base ofensiva, una endonucleasa AP reconoce la desoxirribosa con la base faltante y mella el ADN en ese nucleótido. A continuación, la fosfodiesterasa hidroliza el enlace fosfato-éster restante del fosfato de azúcar 'sin bases', eliminándolo de la cadena de ADN. La ADN polimerasa luego agrega el nucleótido correcto al extremo 3' de la mella. Finalmente, la ADN ligasa III (una versión mamífera dependiente de ATP de la enzima procariota original) sella la mella restante en la cadena. Thomas Lindahl (ver arriba) descubrió la mayoría de estas enzimas.
2. Reparación de escisión de nucleótidos
El descubrimiento de la reparación por escisión de nucleótidos le valió a Aziz Sancar una participación en el Premio Nobel de Química 2015. Los resultados de este mecanismo incluyen la eliminación de dímeros de timidina. Los eventos de reparación por escisión de nucleótidos se muestran a continuación para un dímero de pirimidina.
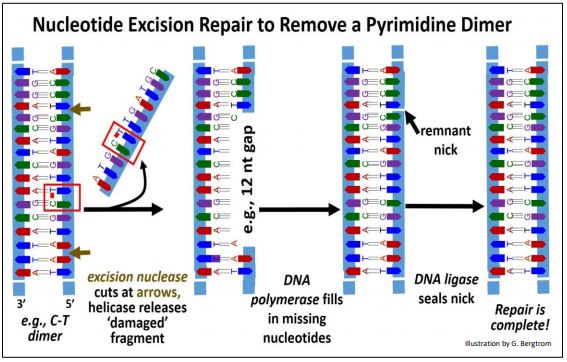
En este ejemplo, una Nucleasa de Escisión reconoce un dímero de pirimidina e hidroliza enlaces fosfodiéster entre nucleótidos a varias bases de distancia de cada lado del dímero. Luego, una helicasa de ADN se desenrolla y separa el fragmento de ADN que contiene las bases dimerizadas de la cadena de ADN dañada. Finalmente, la ADN polimerasa actúa 5'-3' para llenar el hueco y la ADN ligasa sella la mella restante para completar la reparación.
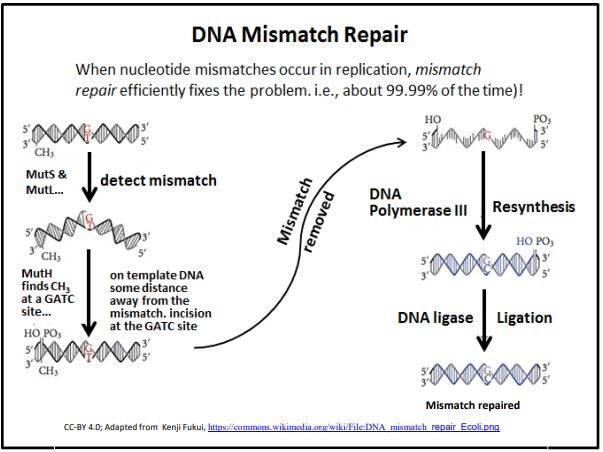
3. Reparación de desajuste
La reparación del desajuste de ADN ocurre cuando la corrección de la ADN polimerasa pierde una inserción incorrecta de la base en una nueva cadena de ADN. Este mecanismo de reparación se basa en el hecho de que el ADN bicatenario muestra un patrón específico de metilación. El descubrimiento del mecanismo de reparación de desajustes le valió a Paul Modrich una participación en el Premio Nobel de Química 2015. Estos patrones de metilación están relacionados con patrones epigenéticos de actividad génica y estructura cromosómica que se espera que sean heredados por las células hijas. Cuando el ADN se replica, los grupos metilo en las cadenas de ADN molde permanecen, pero el ADN recién sintetizado no está metilado. De hecho, tomará algún tiempo para que las enzimas de metilación localicen y metilen los nucleótidos apropiados en el nuevo ADN. En el tiempo intermedio, varias proteínas y enzimas pueden detectar apareamiento de bases inapropiado (los desapareamientos) e iniciar la reparación de desapareamientos. A continuación se ilustra el proceso básico.
4. Reparación Acoplada a la Transcripción (en Eucariotas)
Si una ARN polimerasa que lee un ADN molde encuentra un molde cortado o uno con una sustitución de base inusual, podría interrumpir la transcripción y “no saber qué hacer a continuación”. Por lo tanto, a una pérdida, no se haría una transcripción normal y la célula podría no sobrevivir. No hay gran cosa en un tejido compuesto por miles si no millones de células, ¿verdad? Sin embargo, ¡existe la reparación acoplada de transcripción! En esta vía de reparación, si la ARN polimerasa encuentra una lesión de ADN (es decir, ADN dañado) mientras transcribe una cadena molde, de hecho se detendrá. Esto permite tiempo para que las proteínas de acoplamiento lleguen a la polimerasa estancada y permita que la maquinaria de reparación (por ejemplo, por base o escisión de nucleótidos) efectúe la reparación. Una vez que se completa la reparación, la ARN polimerasa “realiza una copia de seguridad” a lo largo de la cadena molde con la ayuda de otros factores y reanuda la transcripción del molde corregido.
5. Unión de extremos no homólogos
Los errores de replicación del ADN pueden provocar roturas bicatenarias, al igual que los factores ambientales (radiación ionizante, oxidación, etc.). La reparación por unión de extremos no homólogos elimina el ADN dañado y adyacente y vuelve a unir los extremos 'cortados' (se muestran a continuación).
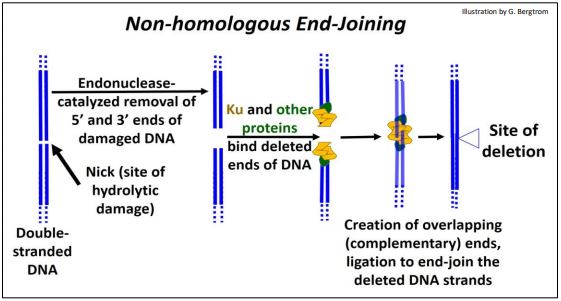
Una vez que se reconoce el sitio de una ruptura bicatenaria, los nucleótidos hidrolizados de los extremos de ambas cadenas en el sitio de ruptura dejan 'extremos romos'. A continuación, varias proteínas (Ku entre otras) unen cadenas de ADN e hidrolizan aún más cadenas de ADN individuales para crear extremos escalonados (superpuestos o complementarios). Los extremos superpuestos de estas cadenas de ADN forman enlaces H. Finalmente, la ADN ligasa sella los extremos superpuestos unidos a H de las cadenas de ADN, dejando una reparación con bases eliminadas.
En las personas mayores, hay evidencia de más de 2000 'huellas' de este tipo de reparación por célula. ¿Cómo es esto posible? Esta reparación rápida a menudo funciona sin efectos nocivos porque la mayor parte del genoma eucariota no codifica genes o incluso ADN regulador (cuyo daño sería de otro modo más grave).
6. Recombinación homóloga
La recombinación homóloga es una parte compleja pero normal y frecuente de la meiosis en eucariotas. Puede recordar que la recombinación homóloga ocurre en la sinapsis en la primera división celular de la meiosis (Meiosis I). Durante la sinapsis, los cromosomas homólogos se alinean. Esto puede conducir a la rotura del ADN, un intercambio de alelos y ligadura para volver a sellar las moléculas de ADN ahora recombinantes. Nuevas recombinaciones de alelos variantes en los cromosomas de espermatozoides y óvulos aseguran la diversidad genética en las especies. El punto clave es que se requiere la rotura del ADN del ADN para intercambiar alelos entre cromosomas homólogos. Consulte el capítulo de genética en un libro de texto introductorio de biología, o el capítulo de recombinación en un texto de genética para recordar estos eventos.
Las células utilizan la misma maquinaria para resellar las roturas de ADN durante la recombinación normal y para reparar el ADN dañado por rotura monocatenaria o bicatenaria. Una sola cadena de ADN mellado durante la replicación puede repararse mediante recombinación con cadenas de ADN homólogo que se están replicando en la otra cadena. Una rotura bicatenaria puede repararse utilizando la misma maquinaria de recombinación que opera en cromátidas hermanas en meiosis. En ambos casos, el proceso repara con precisión el ADN dañado sin ninguna deleción. Estos mecanismos se conservan en las células de todas las especies. Esto indica un imperativo evolutivo de reparación precisa de la supervivencia de las especies, nada menos que el imperativo de mantener la diversidad genética de las especies.
a) Reparación de una Rotura Monocatenaria
Un ejemplo específico de reparación de recombinación homóloga es el restablecimiento de una horquilla de replicación dañada cuando un replisoma alcanza una ruptura en una de las dos cadenas de ADN replicante (ilustrado a continuación).
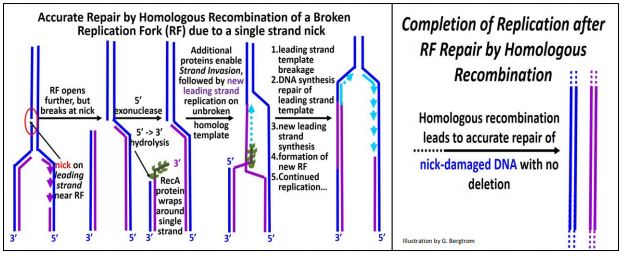
Tal ruptura puede haber ocurrido antes de la replicación misma, y la reparación comienza cuando la horquilla de replicación (RF) alcanza la lesión. En el primer paso, una exonucleasa 5'-3' recorta el ADN molde a lo largo de su complemento recién sintetizado. A continuación, los monómeros de proteína RecA (cada uno con múltiples sitios de unión a ADN) se unen al ADN monocatenario para formar un filamento de nucleoproteína. Con la ayuda de proteínas adicionales, el extremo 3' del 'filamento' escanea la 'otra' cadena replicante en busca de secuencias homólogas. Cuando se encuentran tales secuencias, el filamento de recA-ADN se une a las secuencias homólogas y el filamento de nuevo ADN 'invade' el ADN bicatenario homólogo (es decir, opuesto), separando su molde y ADN recién replicado. Después de la invasión de la cadena, la replicación de una cadena principal continúa desde el extremo 3' de la cadena invasora. Se establece una nueva RF a medida que la plantilla de la cadena principal se rompe y se vuelve a ligar a la rotura original; la nueva replicación de la hebra retrasada se reanuda en la nueva RF (reconstruida). El resultado es una reparación precisa del daño original, sin deleciones o inserciones de ADN.
RecA, una proteína bacteriana, es otra de esas proteínas conservadas evolutivamente. Su homólogo en Archaea se llama RaD. En los eucariotas, el homólogo se llama Rad51, donde inicia la sinapsis durante la meiosis. Así, parece que un papel para RecA y sus homólogos conservados en la reparación del ADN es anterior a su uso en la sinapsis y ¡el cruce en eucariotas! Para más información sobre las funciones de la proteína RecA y sus homólogos, haga clic aquí.
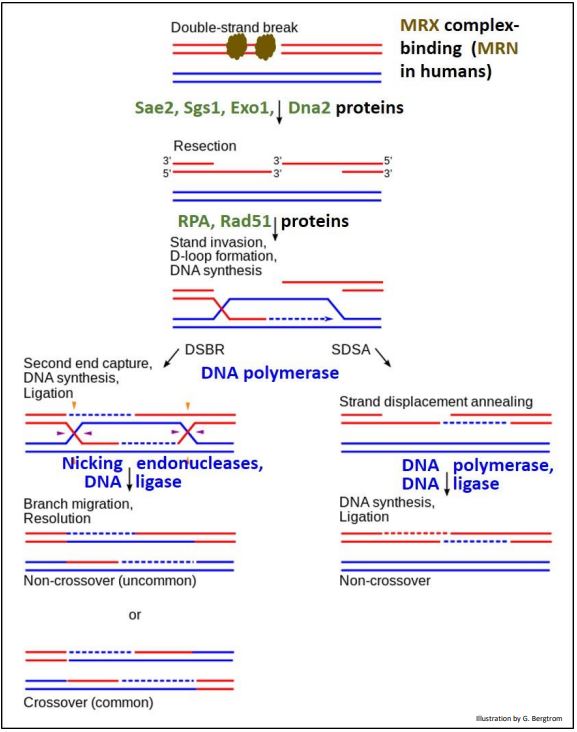
b) Reparación de una Rotura Doble-Trenzada
La recombinación homóloga también puede reparar una rotura de ADN bicatenario con la ayuda de varias enzimas y otras proteínas. Las vías de reparación alternativas se resumen en la ilustración de la página siguiente. Aquí hay una lista de proteínas involucradas en estas vías de recombinación homóloga:
MRX, MRN: unirse a la rotura bicatenaria; reclutar otros factores.
Sae2: una endonucleasa (activa cuando se fosforila).
Ssgs1: una helicasa.
Exo1, Dna2: exonucleasas monocatenarias.
RPA, Rad51, DMC1: proteínas que se unen al ADN saliente para formar un filamento de nucleoproteína e iniciar la invasión de cadenas en secuencias similares.
Se identifican las actividades de otras enzimas en el dibujo. No se muestran en esta ilustración dos productos génicos que interactúan con algunas de las proteínas que median en la vía de reparación. Estos son productos de los genes BRCA1 y BRCA2 (los mismos que al mutar aumentan la probabilidad de que una mujer contraiga cáncer de mama). Expresados principalmente en tejido mamario, sus productos génicos de tipo silvestre (normales) participan en la reparación por recombinación homóloga de roturas de ADN bicatenario. Hacen esto uniéndose a Rad51 (¡el homólogo humano de RecA!).
Cuando se mutan, las proteínas BRCA funcionan mal y el ADN en las células afectadas no se repara de manera eficiente. Esta es la base probable de la mayor probabilidad de contraer cáncer de mama. No ayuda importa que la proteína BRCA1 normal también juegue un papel en la reparación de desapareamientos... ¡y que la proteína mutada no puede! Para terminar este capítulo, ¡aquí hay un poco de ciencia extraña! Lee todo sobre el genoma de una bechicera, casi el 17% de la cual está compuesta por ADN extraño, posiblemente el resultado de aquí.