
11.4: Traducción
- Page ID
- 54084

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)A. Visión general de la traducción (síntesis de proteínas)
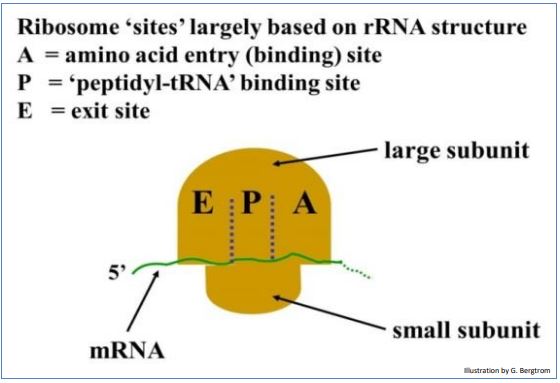
Como cualquier polimerización en una célula, la traducción ocurre en tres etapas: la iniciación une un ribosoma, ARNm y un ARNt iniciador para formar un complejo de iniciación. La elongación es la adición sucesiva de aminoácidos a un polipéptido en crecimiento. La terminación es señalada por secuencias (uno de los codones de parada) en el ARNm y factores de terminación de proteínas que interrumpen la elongación y liberan un polipéptido terminado. Los eventos de traducción ocurren en sitios específicos A, P y E en el ribosoma (ver dibujo a continuación).
B. Traducción - Primeros pasos
1. Elaboración de aminoacil-ARNt
La traducción es quizás el trabajo más intensivo de energía que una célula debe hacer, comenzando con la unión de aminoácidos a sus ARNt. La reacción básica de aminoacilación es la misma para todos los aminoácidos. Una aminoacil-ARNt sintasa específica une cada ARNt a (carga) un aminoácido apropiado. La carga de los ARNt requiere ATP y procede en tres etapas (que se muestran a continuación).
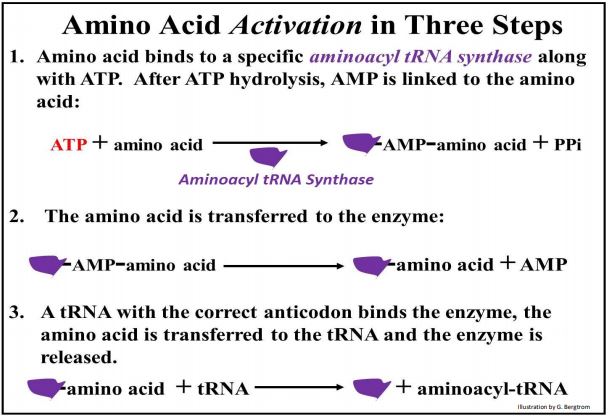
En la primera etapa, ATP y un aminoácido apropiado se unen a la aminoacil-ARNt sintasa. El ATP se hidroliza liberando un pirofosfato (PPi) y dejando un complejo enzima-AMP-aminoácido. A continuación, el aminoácido se transfiere a la enzima, liberando el AMP. Finalmente, el ARNt se une a la enzima, el aminoácido se transfiere al ARNt y la enzima intacta se regenera y libera. El ARNt cargado está listo para su uso en la traducción.
Varios estudios ya habían establecido que los polipéptidos se sintetizan desde su extremo amino (N-) terminal hasta su extremo carboxilo (C-) terminal. Cuando se logró determinar las secuencias de aminoácidos de los polipéptidos, resultó que alrededor del 40% de las proteínas de E. coli tenían una metionina N-terminal, lo que sugiere que todas las proteínas comenzaron con una metionina. También resultó que, aunque solo hay un codón para la metionina, se podrían aislar dos ARNt diferentes para la metionina. Uno de los ARNt se unió a una metionina modificada por formilación, llamada formilmetionina-ARNt fmet (o fMET-ARNt para abreviar). El otro fue metionina-ARNt met (Met-ARNt met para abreviar), cargado con una metionina no modificada.
A continuación se muestran metionina y metionina formilada.
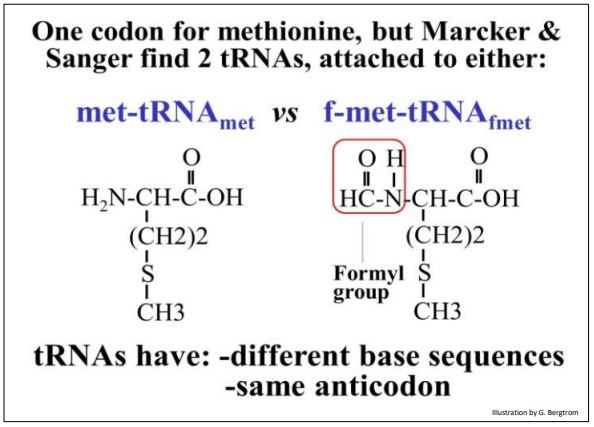
El ARNt met y el ARNt tienen cada uno un anticodón para AUG, el único codón para la metionina, pero tienen diferentes secuencias de bases codificadas por diferentes genes de ARNt. El ARNt met se usa para insertar metionina en el medio de un polipéptido. ARNt es el ARNt iniciador, y es solo utilizado para iniciar nuevos polipéptidos con formilmetionina. En procariotas, la metionina en Met-ARNt se formila en su grupo amino para producir el fMet-ARNf. La enzima formilante que hace esto no reconoce metionina en Met-ARNt met.
En E. coli, una enzima formilasa elimina el grupo formilo de todas las formilmetioninas N-terminales en algún momento después de que la traducción haya comenzado. Como señalamos, la metionina misma (y a veces más aminoácidos N-terminales) también se eliminan de aproximadamente 60% de los polipéptidos de E. coli. Los eucariotas han heredado tanto el ARNf iniciador como el ARNt Met, utilizando solo Met-ARNf durante el inicio. Sin embargo, la metionina en el iniciador eucariota Met-ARNf nunca se formila en primer lugar. Además, la metionina está ausente en prácticamente todos los polipéptidos eucariotas maduros.
Al principio de la evolución, la necesidad de un ARNt iniciador debe haber asegurado un punto de partida correcto para la traducción en un ARNm y por lo tanto el crecimiento de un polipéptido de un extremo a otro, es decir, de su extremo N a su extremo C. En un momento, la formilación de la metionina N-terminal puede haber servido para bloquear la adición accidental de aminoácidos u otras modificaciones en el extremo N-terminal de un polipéptido. Hoy en día, la formilación parece ser una especie de apéndice molecular en bacterias. Desde entonces, la evolución (al menos en eucariotas) ha seleccionado otras características para reemplazar la formilación real como protector del extremo N de polipéptidos.
2. Iniciación
Ahora que hemos cobrado los ARNt, podemos mirar más de cerca los tres pasos de la traducción. La comprensión del inicio de la traducción comenzó con una disección molecular de los componentes de las células de E. coli requeridos para la síntesis de proteínas libres de células (in vitro), incluyendo fraccionamiento celular, purificación de proteínas y experimentos de reconstitución. El inicio comienza cuando la secuencia Shine-Delgarno forma Hbonds con una secuencia complementaria en el ARNr 16S unido a la subunidad ribosómica 30S. La secuencia Shine-Delgarno es una secuencia de nucleótidos corta en la región 5' no traducida (5'-UTR) del ARN mensajero, justo aguas arriba del codón AUG iniciador. Esto requiere la participación de los factores de iniciación IF1 e IF3. En este caso, IF1 e IF3 así como el ARNm se unen a la subunidad ribosómica 30S (abajo).

La demostración de la unión de un ARNm a una subunidad ribosómica requirió el aislamiento y separación de la subunidad ribosómica 30S, una fracción de ARN de la célula y la purificación de proteínas del factor de iniciación de las células bacterianas. Esto fue seguido por la reconstitución (sumar las fracciones separadas de nuevo juntas) en el orden correcto muestran que el ARNm solo se uniría a la subunidad 30S en presencia de las dos proteínas específicas del factor de iniciación.
206 Iniciación de la traducción: mRNA se asocia con la subunidad ribsomal
A continuación, con la ayuda de GTP y otro factor de iniciación (IF2), el iniciador FmettRNAF reconoce y se une al codón de inicio AUG que se encuentra en todos los ARNm. Algunos llaman a la estructura resultante (que se muestra a continuación) el Complejo de Iniciación, que incluye la subunidad ribosómica 30S, Ifs 1, 2 y 3, y el FMET-ARNf.
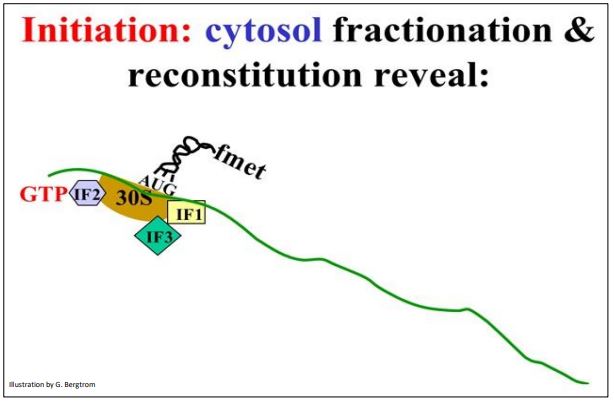
207 Formación del complejo de iniciación
En el último paso de la iniciación, la subunidad ribosómica grande se une a este complejo. Los IFs 1, 2 y 3 se disocian del ribosoma y el iniciador FMET-ARNt fmet termina en el sitio P del ribosoma.
Algunos prefieren llamar a la estructura formada en este punto el complejo de iniciación (abajo).
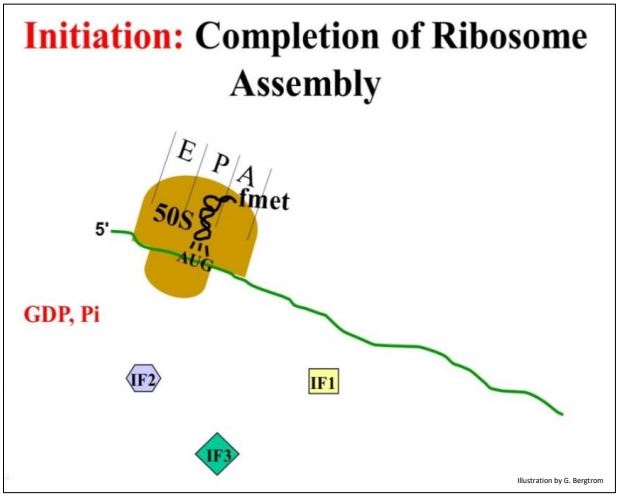
208 Adición de la subunidad ribosómica grande
La iniciación puede ocurrir varias veces en un solo ARNm, formando el polinubosoma, o polisoma descrito en el Capítulo 1. Cada uno de los complejos formados anteriormente se involucrará en la elongación de un polipéptido descrito a continuación.
3. Alargamiento
El alargamiento es una secuencia de reacciones de condensación mediadas por factores proteicos y movimientos de ribosomas a lo largo de un ARNm. Como verá, la elongación de polipéptidos requiere una entrada considerable de energía libre.
a) Alargamiento 1
El primer paso en el alargamiento es la entrada del siguiente aminoacil-ARNt (aa2- ARNtAA2), que requiere la energía libre de hidrólisis de GTP. La energía es suministrada por la hidrólisis del factor de elongación unido a GTP 2 (EF2-GTP). El AA2-ARNtAA2 entra en el ribosoma basándose en la interacción codon-anticodón en el sitio A como se muestra a continuación.
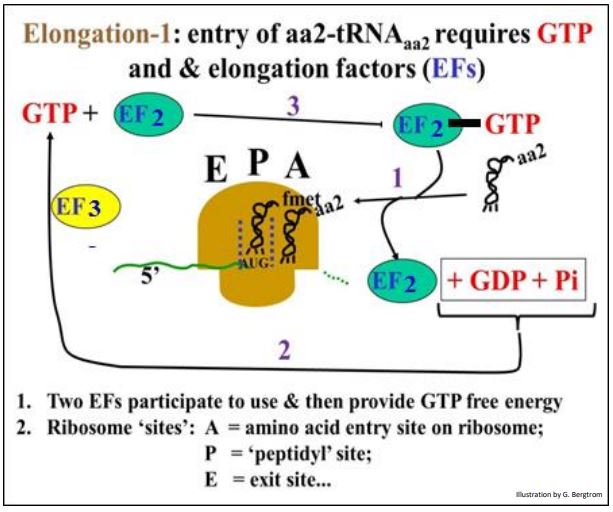
El GDP se disocia de EF2 ya que AA2-ARNtAA2 se une al anticodón en el sitio A. Para mantener el alargamiento en movimiento, el factor de elongación (EF3) refosforila el PIB a GTP, lo que puede volver a asociarse con EF2 libre.
b) Alargamiento 2
La peptidil-transferasa, un componente ribozima del propio ribosoma, une el aminoácido entrante a una cadena en crecimiento en una reacción de condensación.
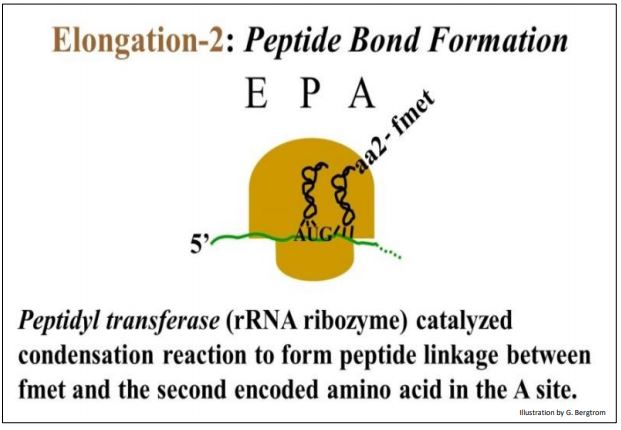
En esta reacción, el fmet se transfiere del iniciador ARNf en el sitio P a AA2-ARNtAA2 en el sitio A, formando un enlace peptídico con aa2.
Elongación 210: una ribozima cataliza la formación de enlaces peptídicos
c) Alargamiento 3
La translocasa cataliza la hidrólisis de GTP a medida que el ribosoma se mueve (se transloca) a lo largo del ARNm. Después de la translocación, el siguiente codón de ARNm aparece en el sitio A del ribosoma y el primer ARNt (en este ejemplo, ARNt) termina en el sitio E del ribosoma.
El movimiento del ribosoma a lo largo del ARNm se ilustra a continuación.
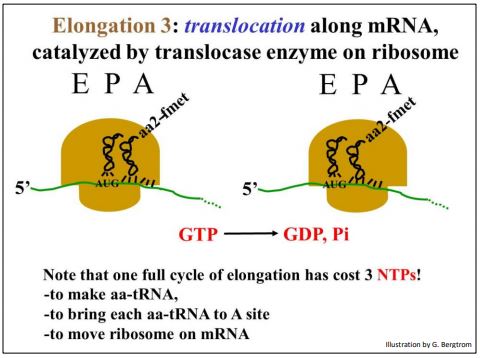
El ARNt, que ya no está unido a un aminoácido, saldrá del sitio E cuando el siguiente (3er) aA-ARNt ingrese al sitio A vacío, basado en una interacción específica codon-anticodón (asistida por factores de elongación y potenciada por hidrólisis GTP) para comenzar otro ciclo de elongación. Obsérvese que en cada ciclo de elongación, se consume un ATP para unir cada aminoácido a su ARNt, y dos GTP se hidrolizan en el ciclo mismo. En otras palabras, a costa de tres NTP, ¡la síntesis de proteínas es la reacción de síntesis de polímeros más cara en las células!
211 Elongación: la translocasa mueve los ribosomas a lo largo del ARNm
212 Adición del Tercer Aminoácido
213 Grandes costos de energía de traducción
A medida que los polipéptidos se alargan, eventualmente emergen de un surco en la subunidad ribosómica grande. Como se señaló, una enzima formilasa en el citoplasma de E. coli elimina el grupo formilo de la fmet de iniciación expuesta de todos los polipéptidos en crecimiento. Mientras que alrededor del 40% de los polipéptidos de E. coli aún comienzan con metionina, proteasas específicas catalizan la eliminación hidrolítica de la metionina amino-terminal (y a veces incluso más aminoácidos) del otro 60% de los polipéptidos. La eliminación del grupo formilo y uno o más aminoácidos N-terminales de nuevos polipéptidos son ejemplos de procesamiento postraduccional.
214 Los destinos de FMet y Met: casos de procesamiento postraduccional
4. Terminación
La traducción de un ARNm por un ribosoma termina cuando la translocación expone uno de los tres codones de parada en el sitio A del ribosoma. Los codones de parada no están situados a cierta distancia del extremo 3' de un ARNm. La región entre un codón de terminación hasta el final del ARNm se denomina región 3' no traducida del ARN mensajero (3'UTR).
Dado que no hay aminoacil-ARNt con un anticodón a los codones de parada (UAA, UAG o UGA), el ribosoma en realidad se detiene y la ralentización de la traducción es lo suficientemente larga para que un factor de terminación proteica ingrese al sitio A. Esta interacción provoca la liberación del nuevo polipéptido y el desmontaje de las subunidades ribosómicas del ARNm. El proceso requiere energía de otra hidrólisis GTP. Después de la disociación, las subunidades ribosómicas se pueden volver a ensamblar con un ARNm para otra ronda de síntesis de proteínas. La terminación de la traducción se ilustra a continuación.
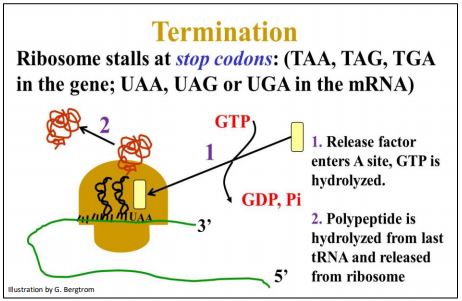
215 Terminación de la traducción
Hemos visto algunos ejemplos de procesamiento postraduccional (eliminación de grupos formilo en E. coli, eliminación de la metionina N-terminal de la mayoría de polipéptidos, etc.) La mayoría de las proteínas, especialmente en eucariotas, se someten a una o más etapas adicionales de procesamiento postraduccional antes de convertirse en biológicamente activas. Veremos ejemplos en los próximos capítulos.
¡Concluyamos este capítulo con un momento de “pensábamos que lo sabíamos todo”! Un estudio reciente informa que los ribosomas a veces pueden reiniciar la traducción en la UTR 3' de un ARNm usando codones AUG aguas arriba del codón de inicio normal del ARNm. ¡Hay evidencia de que los polipéptidos cortos resultantes pueden ser funcionales! Da click aquí para leer más: aquí.