13.3: Regulación eucariota de la traducción
- Page ID
- 54566

A. Fundamentos de la Traducción de ARNm eucariota
A continuación se muestran las características básicas del inicio de la traducción en eucariotas.
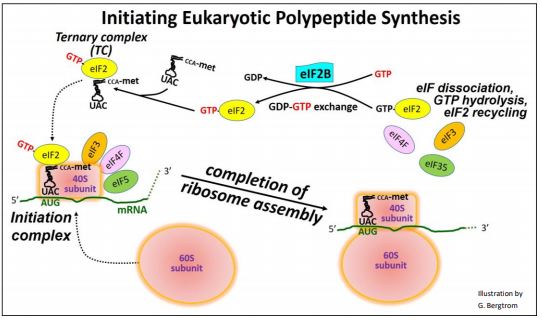
En muchos aspectos, el proceso general es similar al inicio de la traducción procariota descrito en otra parte. La subunidad ribosómica 40S en sí misma puede unirse y escanear un ARNm, buscando el sitio de inicio de un ORF (marco de lectura abierto) que codifica un polipéptido. Cuando el factor de iniciación eucariota 2 unido a GTP (GTP-eIF2) se une a Met-ARNf, forma un complejo ternario (TC). El TC puede asociarse con la subunidad 40S de barrido. Cuando una subunidad de exploración asociada a TC se encuentra con el sitio de inicio del ORF, la exploración se detiene. Los EIF adicionales permiten la formación del complejo de iniciación, posicionando el anticodón de ARNt iniciador sobre el sitio de inicio AUG en el ARNm. El complejo de iniciación luego recluta la subunidad ribosómica grande (60S). La unión de la subunidad ribosómica 60S al complejo de iniciación provoca la liberación de todos los EIF y la hidrólisis del GTP en eIF2. El PIB se mantiene vinculado a eIF2. Para que las síntesis de proteínas continúen, el nuevo GTP debe reemplazar al PIB en eIF2. Otro factor de iniciación, eIF2b, facilita esta permuta GTP/PIB, reciclando GTP-EIF2 para su uso en iniciación. La regulación de la traducción se superpone a estos procesos básicos.
B. Reglamento de traducción
Ya que los ARNm están hechos para ser traducidos, es probable que por defecto, ¡lo sean! Sabemos que las colas CAP y poli (A) en los ARNm son necesarias para una traducción eficiente porque los ARNm diseñados para carecer de uno y/o el otro están mal traducidos. Además, hay poca evidencia de que las células modifiquen el proceso de caperuza o poliadenilación, o las propias estructuras.
La regulación de la traducción generalmente apunta al inicio. Puede ser global, afectando la síntesis de muchos polipéptidos a la vez, o específica, afectando a un solo polipéptido. La regulación global implica cambios en la actividad de los factores de iniciación eucariotas (EIF) que típicamente afectarían a toda la síntesis de proteínas celulares. La regulación específica implica la unión de secuencias o regiones en uno o unos pocos ARNm que reconocen y se unen a proteínas reguladoras específicas y/u otras moléculas. Esa unión controla la traducción de solo esos ARNm, sin afectar la biosíntesis general de proteínas. Las características estructurales del ARNm involucradas en la traducción y en la regulación de la traducción se ilustran a continuación.
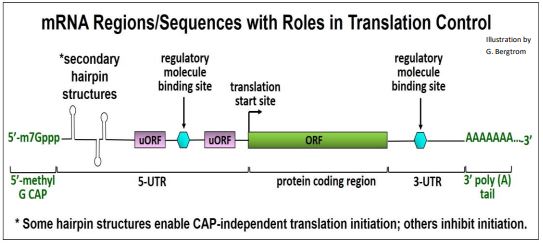
Consideraremos tres ejemplos de control traduccional de la expresión génica.
1. Control específico de la traducción mediante proteínas de unión a ARNm
La ferritina es una proteína celular de almacenamiento de hierro compuesta por polipéptidos de cadena pesada y ligera. Se inhibe la traducción de ferritina en células deficientes de hierro. En ausencia de producción de ferritina, los complejos ferritina-hierro liberan hierro para uso metabólico. La 5'-UTR de los ARNm para ambas cadenas contiene sitios de unión tallo-bucle que reconocen específicamente proteínas reguladoras de hierro (IRP1, IRP2). Cuando los ARNm de ferritina se unen a IRP, se bloquea el inicio de la traducción. La inhibición de la traducción de ferritina por IRP se ilustra a continuación.
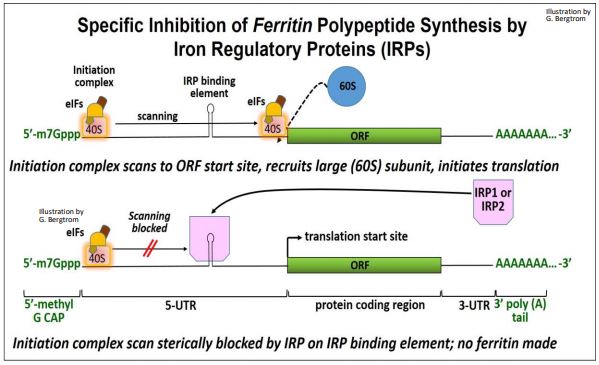
Normalmente, el complejo de iniciación escanea la 5'-UTR de un ARNm. Cuando encuentra el sitio de inicio de la traducción normal, puede unirse a la subunidad grande y comenzar a traducir el polipéptido. En las células deficientes de hierro, se cree que el escaneo por el complejo de iniciación está físicamente bloqueado por obstáculos estéricos.
2. Coordinación de la síntesis de hemo y globina
Considera que los reticulocitos (los precursores de los eritrocitos, los glóbulos rojos en los mamíferos) sintetizan proteínas globinas. También sintetizan hemo, una molécula de anillo de porfirina unida a hierro. Cada globina debe unirse a un solo hemo para formar una subunidad de proteína de hemoglobina. Claramente, no serviría para que un reticulocito produjera demasiada proteína globina y no suficiente hemo, o viceversa. Resulta que la hemina (un precursor del hemo) regula el inicio de la traducción tanto\(\alpha \) de ARNm como\(\beta \) de globina. Recordemos que, para sostener la traducción del ARNm de globina, el GDP-eIF2 generado después de cada ciclo de elongación de la traducción debe ser intercambiado por GTP fresco. Esto se ve facilitado por el factor de iniciación eIF2b. eIF2b puede existir en estados fosforilados (inactivos) o no fosforilados (activos). Asegurarse de que la globina no esté subproducida o sobreproducida en relación con la biosíntesis de hemo implica controlar los niveles de eIF2b activo frente a inactivo por hemina. La hemina se acumula cuando no hay suficiente polipéptido de globina para combinarse con el hemo en la célula. El exceso de hemina se une e inactiva a una HCR quinasa, evitando la fosforilación de eIF2b. Dado que el eIF2b no fosforilado está activo, facilita la permuta GTP/PIB necesaria para permitir la traducción continua. Por lo tanto, la iniciación continua asegura que la traducción de ARNm de globina pueda mantenerse al día con los niveles de hemo En otras palabras, si la producción de hemina se adelanta a la globina, promoverá más traducción de globina.
Cuando los niveles de globina y hemo se vuelven aproximadamente equimolares, la hemina ya no está en exceso. Luego se disocia de la quinasa HCR activa. La quinasa ahora activa cataliza la fosforilación de eIF2b. El fosfo-eIF2B está inactivo y no puede facilitar la permuta GTP/PIB en eIF2. El inicio de la traducción de ARNm de globina, así bloqueado, permite una menor tasa de traducción de polipéptidos de globina para mantener el ritmo de la síntesis de hemo. La regulación del inicio de la traducción de ARNm de globina por hemina se muestra a continuación.
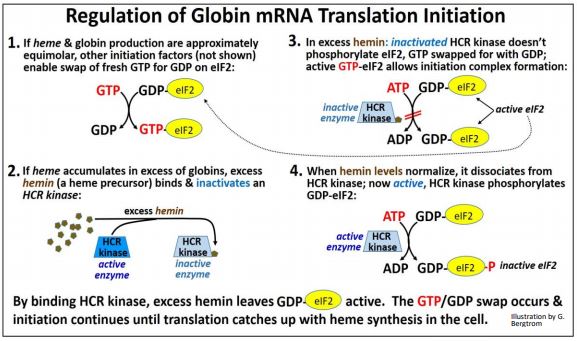
237 Regulación de traducción de la síntesis de polipéptidos de globina
3. Regulación Traslacional de Levadura GCN4
Al igual que la coordinación de la producción de hemo y globina, la regulación de la proteína GCN4 se basa en controlar la capacidad de las células para intercambiar GTP por PIB en eIF2. Sin embargo, esta regulación es bastante más compleja, ¡a pesar de que la levadura es un eucariota más primitivo! ¡GCN4 es un factor de transcripción global que controla la transcripción de hasta 30 genes en vías para la síntesis de 19 de los 20 aminoácidos! El descubrimiento de que la inanición de aminoácidos provocó que las células de levadura aumentaran su producción de aminoácidos en las células condujo al descubrimiento del mecanismo General de Control de Aminoácidos (GAAC) que involucra a GCN4. GCN es la abreviatura de G eneral C ontrol N ondepressible, refiriéndose a sus efectos regulatorios globales positivos. Resulta que la proteína GCN4 también está involucrada en la expresión génica del estrés, la homeostasis del glucógeno, la biosíntesis de purinas..., ¡de hecho en la acción de hasta el 10% de todos los genes de levadura! Aquí nos enfocamos en el mecanismo GAAC.
Las células de levadura provistas de amplios aminoácidos no necesitan sintetizarlas. Bajo estas condiciones, el GCN4 está presente en niveles basales (es decir, bajos). Cuando las células están privadas de aminoácidos, los niveles de GCN4 aumentan hasta diez veces en dos horas, lo que resulta en un aumento en la síntesis general de aminoácidos. Esta respuesta rápida se produce porque la inanición de aminoácidos señala un aumento en la actividad de GCN2, una proteína quinasa. La quinasa GCN2 cataliza la fosforilación de GdPEIF2. Como ya hemos visto, el eIF2b fosforilado no puede intercambiar GTP por PIB en el eIF2, en este caso con los resultados que se muestran a continuación.
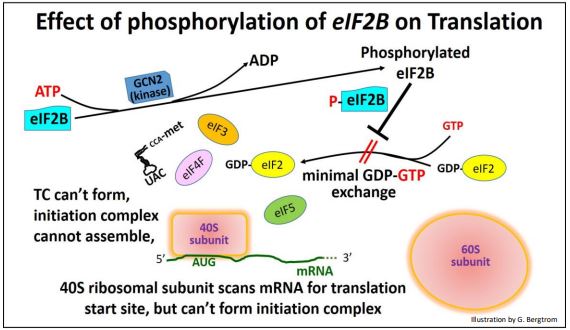
Aquí hay una paradoja. Se esperaría una desaceleración en la regeneración de GTP-eIF2 para inhibir la síntesis general de proteínas, y lo hace. Sin embargo, los niveles reducidos de GTP-eIF2 de alguna manera también estimulan la traducción del ARNm de GCN4, lo que lleva a una mayor transcripción de los genes de síntesis de aminoácidos. En otras palabras, la inanición de aminoácidos lleva a las células de levadura a utilizar sustratos disponibles para elaborar sus propios aminoácidos con el fin de que la síntesis de proteínas pueda continuar... ¡al mismo tiempo que se desactiva la formación del complejo de iniciación!
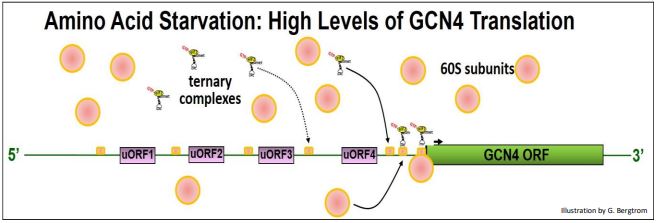
Aceptemos esa paradoja por ahora, y veamos cómo la inanición de aminoácidos conduce a una mayor traducción de la proteína GCN4 y a la sobrerregulación de las vías de biosíntesis de aminoácidos. Para empezar, vamos a necesitar entender la estructura del ARNm de GCN4. En la siguiente ilustración, anote los 4 UORF cortos en la 5'UTR del ARN; estos juegan un papel clave en la regulación de la traducción GCN4.
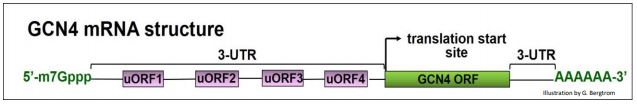
Señalamos anteriormente que cuando una subunidad ribosómica 40S asociada a un complejo ternario (TC) escanea un ARNm y encuentra los sitios de inicio de ORF para su polipéptido, se forman complejos de iniciación, 60 subunidades ribosómicas se unen y comienza la traducción. El ARNm de GCN4 tiene cuatro UORF en su 5' UTR. Si bien los UORF codifican solo unos pocos aminoácidos antes de encontrar un codón de parada, también pueden reconocerse durante el escaneo. Cuando las TCs y las subunidades 40S son abundantes, parecen involucrar UORF en preferencia al ORF de la región de codificación GCN4, como se ilustra a continuación.
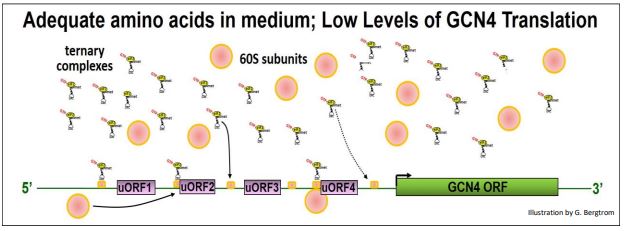
En estas condiciones, el eIF2b activo permite la permuta GTP/PIB sobre el PID-EIF2, lo que lleva a un reciclaje eficiente de GTP-eIF2 y altos niveles de TC. Los TC se unen a pequeñas subunidades durante el escaneo y/o en los sitios de inicio de los UORF, formando complejos de iniciación que luego se unen a subunidades ribosómicas 60S y comienzan la traducción de UORF. El efecto es ralentizar el barrido más allá de los UORF, inhibiendo así la formación del complejo de iniciación en el ORF real de GCN4.
¿Qué sucede en cultivos de células de levadura privados de aminoácidos, cuando GTP-eIF2 no se puede regenerar de manera eficiente y los TCs escasean? Para revisar, la inanición de aminoácidos señala un aumento en la actividad de la quinasa GCN2 que resulta en la fosforilación e inactivación de eIF2b. La fosfo-eIF2 inactiva no facilitará la permuta GTP/PIB en GDP-eIF2, inhibiendo la síntesis global de proteínas. La reducción resultante en GTP-eIF2 también disminuye los niveles de TC y subunidades 40S asociadas a TC. La siguiente ilustración muestra cómo este fenómeno regula positivamente la traducción de GCN4, incluso cuando la traducción de otros ARNm ha disminuido.
C. Regulación del volumen de proteínas (vida media)
Ya hemos visto que los orgánulos tienen una vida útil finita, o semivida. Recordemos que los lisosomas participan en la destrucción de mitocondrias desgastadas y sus componentes moleculares. También vimos el papel de los ARN pequeños (especialmente miARN) en la destrucción de ARN viejos, dañados o no deseados de las células. Todas las estructuras celulares y moléculas tienen una vida media finita, definida como el tiempo que tarda la mitad de ellas en desaparecer en ausencia de una nueva síntesis de la estructura o molécula. Como ya sabemos, el nivel de estado estacionario de cualquier estructura celular o molécula existe cuando la velocidad de su fabricación o síntesis se equilibra con la tasa de su recambio. Por supuesto, los niveles de estado estacionario de las cosas pueden cambiar. Por ejemplo, el nivel de expresión génica (la cantidad de un ARN final o producto génico proteico en una célula) puede cambiar si cambian las tasas de transcripción, procesamiento o recambio. También debemos esperar lo mismo para los niveles de estado estacionario de las proteínas celulares. Aquí consideramos los factores que rigen la vida media de las proteínas celulares.
La vida media de diferentes proteínas parece ser inherente a su estructura. Así, algunas cadenas laterales de aminoácidos están más expuestas en la superficie de la proteína y, por lo tanto, son más susceptibles de cambiar o dañar con el tiempo que otras. Las proteínas con menos aminoácidos 'vulnerables' deben tener una vida media más larga que las que tienen más de ellos. Las proteínas dañadas por errores de traducción, plegado, procesamiento que salió mal o simplemente desgastadas por el uso o la 'vejez” serán atacadas para su destrucción. ¡Todas las moléculas tienen una vida media!
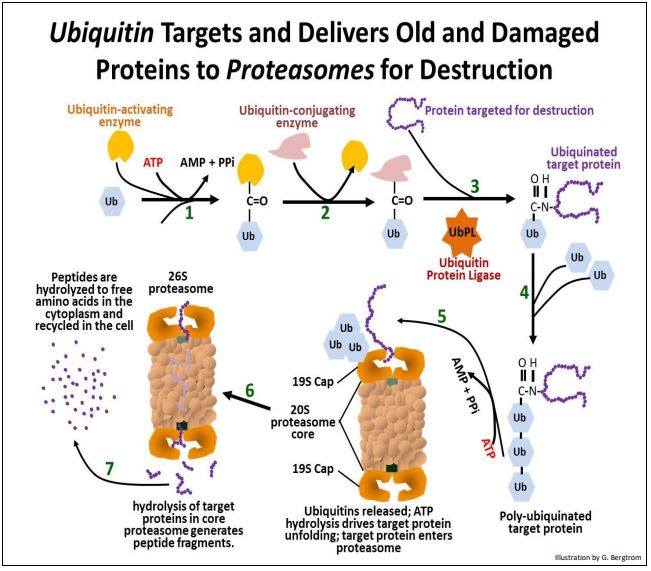
El mecanismo para detectar y destruir proteínas indeseadas viejas, dañadas o mal engendradas involucra un polipéptido de 76 aminoácidos llamado ubiquitina que se dirige a la proteína para su destrucción, entregándola a un gran complejo de polipéptidos llamado el proteasoma. Esto es lo que sucede:
- El primer paso es activar una ubiquitina. Esto comienza cuando la hidrólisis de ATP alimenta la unión de ubiquitina a una enzima activadora de ubiquitina.
- Luego, una enzima conjugadora de ubiquitina reemplaza a la enzima de activación de ubiquitina.
- La proteína destinada a la destrucción reemplaza a la enzima conjugadora de ubiquitina.
- Varias ubiquitinas más luego se unen a este complejo.
- La proteína poli-ubiquinada entrega su proteína a una de las estructuras 19S 'CAP' de un proteasoma.
- Después de unirse a una de las estructuras CAP de un proteasoma, las proteínas diana poli-ubiquinadas se disocian y las ubiquitinas se liberan y reciclan a medida que la proteína diana se despliega (alimentada por hidrólisis de ATP). La proteína desplegada luego ingresa a un proteasoma de núcleo 20S.
La proteína diana es digerida a fragmentos peptídicos cortos por enzimas proteolíticas en el interior del núcleo del proteasoma. Los fragmentos se liberan del complejo CAP en el otro extremo del proteasoma y se digieren hasta aminoácidos libres en el citoplasma. Hay una variedad alucinante de proteínas en una célula..., y hay hasta 600 proteínas de ubiquitina diferentes, ¡codificadas por tantos genes! Presumiblemente, cada ubiquitina maneja una subclase de proteínas basadas en características comunes de su estructura.
Con su compleja estructura cuaternaria, el proteasoma 26S es más pequeño que una subunidad ribosómica pequeña eucariota (40S), pero sigue siendo una de las partículas citoplásmicas más grandes... ¡y sin el beneficio de ningún ARN en su estructura! La ilustración de la siguiente página detalla el papel de la ubiquitina en la degradación de una proteína desgastada por un proteasoma. Haz clic en Proteasoma en Acción para ver una versión animada de la ilustración.