
13.2: Control postranscripcional de la expresión génica
- Page ID
- 54582

No hace mucho pensábamos que muy poco del genoma eucariota había sido transcrito alguna vez. También pensamos que los únicos ARN no codificantes eran ARNt y ARNr. Ahora sabemos que otros ARN juegan un papel en la regulación génica y la degradación del ADN celular gastado o ADN extraño no deseado. Estos se discuten en detalle a continuación.
A. Ribosomas
Los ribointerruptores son un mecanismo de transcripción bacteriana para regular la expresión génica. Si bien este mecanismo no es específicamente postranscripcional, se incluye aquí porque la acción ocurre después del inicio de la transcripción y aborta la finalización de un ARNm. Cuando se transcribe el ARNm de una enzima en la vía de síntesis de guanina, se pliega en estructuras de tallo y bucle. La síntesis enzimática continuará durante el tiempo que la célula necesite para producir guanina. Pero si la guanina se acumula en la célula, el exceso de guanina se unirá a elementos tallo-bucle cerca del extremo 5' del ARNm, haciendo que la ARN polimerasa y el ARNm parcialmente completado se disocien del ADN, finalizando prematuramente la transcripción. La base de la regulación de guanina riboswitch de la expresión de una enzima de la vía de síntesis de guanina se muestra a continuación.
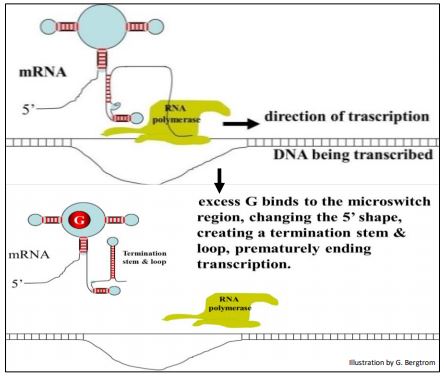
232 riboswitches interrumpen la transcripción bacteriana
La capacidad de formar estructuras de tallo-bucle plegadas en los extremos 5' de los ARNm bacterianos parece haber permitido la evolución de las estrategias de regulación de la traducción. Mientras que la interacción de guanina con la estructura tallo-bucle de un ARNm 5' emergente puede abortar su propia transcripción, interacciones similares de metabolito/ARNm pequeños e incluso proteína/ARNm también pueden regular (en este caso prevenir) la traducción. Como veremos en breve, las estructuras plegadas de ARNm 5' también juegan un papel en la regulación de la traducción eucariota.
233 pequeños metabolitos también regulan la traducción de ARNm bacteriano
B. CRISPR/Cas: Complejo ARN-proteína de un sistema inmune adaptativo procariota
En organismos superiores, el sistema inmune es adaptativo. Se acuerda de la exposición previa a un patógeno, y así puede montar una respuesta a una segunda exposición al mismo patógeno. El descubrimiento de un 'sistema inmune adaptativo' en muchos procariotas (bacterias, arquebacterias) fue, por lo tanto, una especie de sorpresa.
Los ARN de CRISPR (C lustered R egularly I nterspaced S hort P alindrómico R epeat) se derivan de transcritos de fagos que han interactuado con proteínas asociadas a CRISPR-asociadas (Cas). Conforman el sistema CRISPR/ Cas que parece haber evolucionado para combatir la infección viral al apuntar el ADN del fago para su destrucción. Cuando el ADN viral ingresa a una célula durante una infección fágica, puede generar una matriz de genes CRISPR/Cas en el genoma bacteriano, con secuencias de ADN espaciadoras que separan las repeticiones de los genes CRISPR. Estos remanentes de una infección fágica son la memoria de este sistema inmune p rocariota. Cuando un fago intenta reinfectar una célula previamente expuesta, se transcriben ARN espaciadores y genes Cas. Después de la traducción del ARNm de Cas, la proteína Cas y los ARN espaciadores se acoplarán y se dirigirán al ADN del fago entrante para su destrucción para prevenir Así, los sistemas CRISPR/Cas (¡hay más de uno!) recordar ataques de fagos anteriores y transmitir esa memoria a las células de la progenie. El sistema CRISPR/Cas9 en Streptococcus pyogenes es uno de los más simples de estos sistemas de defensa inmune (ilustrado a continuación).
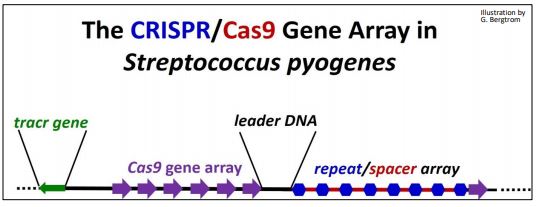
La matriz de genes CRISPR/Cas consta de los siguientes componentes:
- Cas: Genes nativos de células hospedadoras
- CRISPR: repeticiones de 24-48 pb nativas de células hospedadoras
- ADN espaciador: ADN entre repeticiones CRISPR: típicamente, ADN de fago de infección previa de fagos o transformación plasmídica
- ADN líder: Contiene promotor para la transcripción de ARN CRISPR/espaciador
- gen tracr: Codifica el ARN activador de la transcripción (tracr) (no todos los sistemas)
Echemos un vistazo a CRISPR/Cas en acción.
1. La respuesta inmune CRISPR/Cas
Considerar el mecanismo de acción de este sistema inmune procariota. La acción comienza cuando el ADN infeccioso del fago entra en la célula, como se dibuja a continuación.
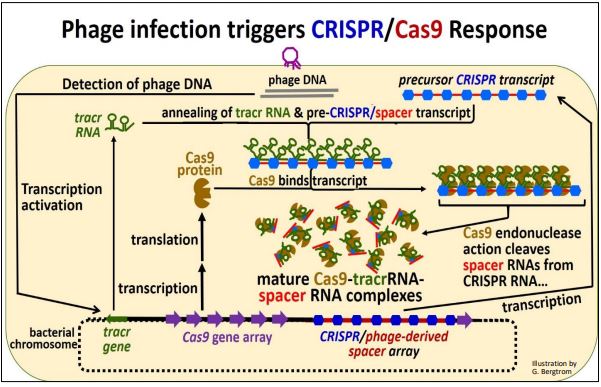
Resumimos lo que ha sucedido aquí:
a) El ADN del fago entrante se detectó después de la infección del fago.
b) Luego se transcriben los genes tracr y Cas junto con la región CRISPR/Spacer. Los ARNm Cas se traducen para producir la proteína Cas. Recuerde, los ADN espaciadores en la región CRISPR son el legado de una infección previa por fagos.
c) El ARN CRISPR/espaciador forma enlaces de hidrógeno con una región complementaria del ARN tracr ya que los dos ARN se asocian con proteínas Cas.
d) Las endonucelasas de proteína Cas hidrolizan ARN espaciador de secuencias de ARN CRISPR. Los ARN espaciadores permanecen asociados con el complejo mientras que las secuencias CRISPR palindrómicas reales imperfectamente palindrómicas (mostradas en azul en la ilustración anterior) se caen.
En los siguientes pasos, los ARN espaciadores derivados de fagos, ahora llamados ARN guía (o ARNg), 'guían' complejos de Cas9/tracrRNA/ARN espaciador maduros a nuevo ADN de fago entrante resultante de un ataque de fago. La asociación del complejo con el ADN del fago entrante y los eventos posteriores se ilustran a continuación.
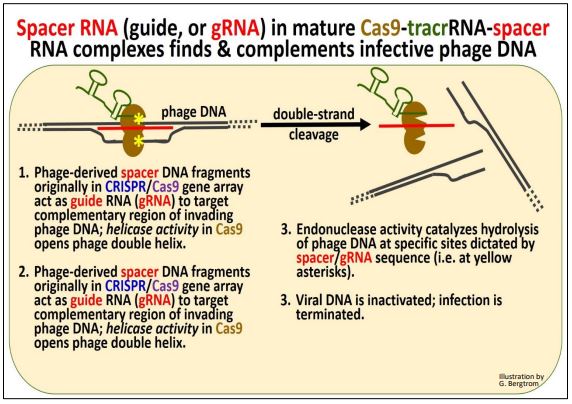
Una vez más, resumimos:
a) El espaciador (es decir, ARNg) en el complejo se dirige al ADN del fago entrante.
b) La helicasa Cas desenrolla el ADN del fago entrante en regiones complementarias.
c) El ARNg se une al ADN del fago entrante.
d) Las endonucleasas Cas crean una rotura bicatenaria (escisión hidrolítica) en sitios específicos en el ADN del fago entrante. Debido a que la escisión de la cadena de ADN del sitio preciso está guiada por moléculas de ARN, las endonucleasas CRISPR/Cas se clasifican como enzimas de restricción tipo V.
e) Se destruye el ADN del fago entrante y se aborta una nueva infección por fagos.
Consulte aquí para obtener más información sobre cómo las bacterias adquieren ADN espaciadores y, por lo tanto, cómo este sistema inmunitario adaptativo primitivo 'recuerda') en primer lugar
2. Uso de CRISPR/Cas para Editar/Ingeniería de Genes
Los primeros estudios demostraron la escisión reproducible del ADN del fago entrante en nucleótidos específicos. ¡Varios laboratorios se dieron cuenta rápidamente de que podría ser posible adaptar el sistema para cortar ADN en prácticamente cualquier nucleótido específico en un ADN diana! Ha resultado que el sistema funciona tanto in vivo como in vitro, permitiendo un potencial prácticamente ilimitado para editar genes y ARN en un tubo de ensayo... o en cualquier célula. Aquí está el proceso básico:
a) Diseñar ADNg con una secuencia de ADN específica de Cas que se dirige a una diana deseada en el ADN genómico.
b) Fusionar el ADNg con el ADN tracr para hacer un solo ADN guía (ADNgs) de manera que se pueda hacer como una única transcripción guía (ARNgs).
c) Diseñar una matriz de genes CRISPR/Cas9 que sustituya este ADNgs por sus ADN espaciadores originales.
d) Colocar matriz modificada por ingeniería genética en un plásmido junto a promotores regulados.
e) Transformar células por 'electroporación' (¡funciona para casi cualquier tipo de célula!)
f) Activar el promotor para transcribir los genes CRISPR/Cas9...
Las aplicaciones son poderosas... ¡y polémicas!
3. El poder y la polémica
La aplicación de la edición génica con sistemas CRISPR/Cas ya ha facilitado estudios de la función génica in vitro, en células y en organismos enteros. Haga clic aquí para obtener una descripción de las aplicaciones CRISPR/Cas que ya están en el mercado! La eficiencia de la edición génica específica usando sistemas CRISPR/Cas es muy prometedora para comprender la estructura y función básica del gen, para determinar la base genética de la enfermedad y para acelerar la búsqueda de terapias génicas. Estos son solo algunos ejemplos de cómo se están aplicando los enfoques CRISPR/Cas.
- Se puede diseñar un ARNgs con mutaciones deseadas que se dirigen a sitios específicos en el ADN cromosómico. Luego clona ARNgs en la matriz CRISPR/Cas9 en un plásmido. Después de la transformación de las células apropiadas, el CRISPR/Cas9 diseñado forma un complejo con secuencias de ADN diana. Después del corte de ambas cadenas del ADN diana, la reparación del ADN puede insertar las secuencias guía mutadas en el ADN diana. El resultado es la pérdida o adquisición de secuencias de ADN en sitios específicos, exactos o Precision Gene Editing. Es la capacidad de hacer esto en células vivas lo que ha excitado a las comunidades de investigación básica y clínica.
- Antes de transformar las células, diseñe la matriz de genes CRISPR/Cas9 en el plásmido para eliminar ambas actividades endonucleasas de la proteína Cas. Tras la transcripción de la matriz en células transformadas, el CRISPR/Cas9-sgRNA todavía encuentra un gen dirigido a ARNgs. Sin embargo, al carecer de actividades de la proteína endonucleasa CAS, el complejo que se forma simplemente se asienta ahí bloqueando A esta técnica se le denomina a veces CRISPRI (interferencia CRISPER), por analogía con la ARNi. Aplicado a organismos (y no solo in vitro o a células), imita los experimentos de mutación knockout mucho más difíciles que se han utilizado en estudios de comportamiento de células u organismos que se vuelven incapaces de expresar una proteína específica.
- Ahora hay varios sistemas CRISPR/Cas en funcionamiento capaces de Edición Genética de Precisión. Son emocionantes por su velocidad, precisión, sus perspectivas de terapias génicas rápidas y dirigidas para combatir enfermedades y sus posibilidades de alterar poblaciones enteras (llamadas Gene Drive). Al insertar genes modificados en las células de la línea germinal de los organismos diana, el impulso génico puede hacer inofensivas poblaciones enteras de mosquitos de la paludismo, para eliminar la resistencia a pesticidas en, por ejemplo, insectos, eliminar la resistencia a herbicidas en plantas indeseables o eliminar genéticamente especies invasoras. Para obtener más información, haga clic en Impulsión génica; para una lectura fácil sobre este proceso y las controversias que rodean las aplicaciones de las tecnologías CRISPR a los mosquitos en particular, consulte J. Adler, (2016) Un mundo sin mosquitos. Smithsonian, 47 (3) 36-42, 84.
- Incluso es posible eliminar un cromosoma completo de las células. Este poco de ingeniería genética global se basa en identificar múltiples secuencias únicas en un solo cromosoma y luego dirigirse a estos sitios para CRISPR/Cas. Cuando se activa el sistema, el cromosoma se corta en esos sitios, fragmentándolo más allá de la capacidad de los mecanismos de reparación del ADN para arreglar la situación. Haga clic aquí para obtener más información.
Si no por otra razón que su eficiencia y simplicidad, la edición de genes de precisión con técnicas CRISPR/Cas ha planteado cuestiones éticas. Claramente, existe el potencial para el abuso, o incluso para su uso sin ningún propósito beneficioso en absoluto. Es significativo que, como en todas las discusiones sobre ética biológica, los científicos estén muy comprometidos en la conversación. A pesar de la polémica, sin duda seguiremos editando genes con CRISPR/Cas, ¡y podremos buscar un futuro cercano Premio Nobel por su descubrimiento y aplicación! Si aún tienes reparos, tal vez la edición de ARN sea la respuesta. Consulta el enlace en ¿Por qué editar ARN? para una visión general de las posibilidades!
Finalmente, “ratones y hombres” (y mujeres y bebés también) tienen anticuerpos contra las proteínas Cas9, lo que sugiere una exposición previa a antígenos CRISPR/Cas9 microbianos. ¡Esta observación puede limitar las aplicaciones clínicas de la tecnología! Ver Futuro Incierto de la Tecnología CRISPR-Cas9.
C. Los ARN pequeños: miARN y ARNi en eucariotas
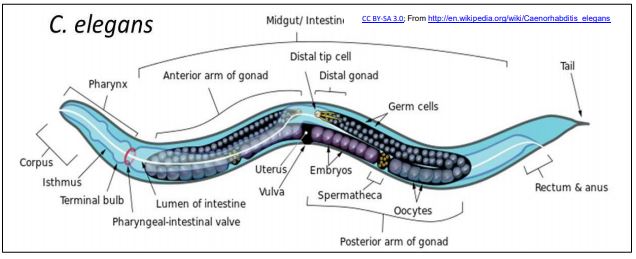
Los micro ARN (miRNAs) y los ARN interferentes pequeños (ARNip) se encuentran en C. elegans, un pequeño nematodo (gusano redondeado) que rápidamente se convirtió en un modelo para estudios de biología y desarrollo celular y molecular. Los atractivos particulares de C. elegans son que (a) su genoma tiene ~21,700 genes, ¡comparables a los ~25,000 genes en un genoma humano! ; b) utiliza los productos de estos genes para producir un gusano adulto que consiste en apenas 1031 células organizadas en todos los órganos principales que se encuentran en organismos superiores; c) ¡Es posible rastrear los orígenes embrionarios de cada célula en su cuerpo! C. elegans se ilustra a continuación.
1. ARN interferente pequeño (siRNA)
El ARNIp se encontró por primera vez tanto en plantas como en C. elegans. Sin embargo, los ARNip (y miRNAs) son comunes en muchos organismos superiores; los ARNip fueron llamados así porque interfieren con la función de otros ARN extraños a la célula u organismo. Su acción se denominó interferencia de ARN (ARNi). Por su descubrimiento de los ARNip, A. Z. Fire y C. C. Mello compartieron el Premio Nobel de Fisiología o Medicina 2006. La acción del ARNIp dirigido a ADN extraño se ilustra a continuación.
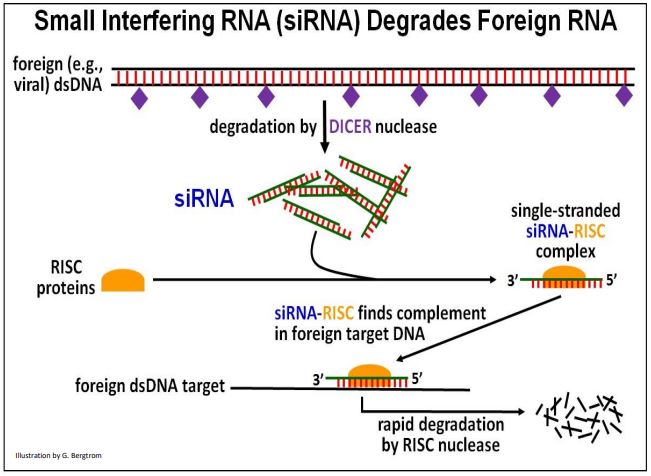
Cuando las células reconocen ARN bicatenarios extraños (por ejemplo, algunos genomas de ARN virales) como extraños, el DICER una nucleasa llamada los hidroliza. Los productos de hidrólisis bicatenarios cortos resultantes (los ARNip) se combinan con R NaI I nduced S ilering C omplex, o proteínas RISC. La cadena de ARNip antisentido en el complejo ARNsi-RISC resultante se une a regiones complementarias de ARN foráneos, dirigiéndolos para su degradación. El uso celular de RISC para controlar la expresión génica de esta manera puede haber derivado del uso de proteínas RISC por los miARN como parte de un mecanismo de defensa celular, que se discutirá a continuación.
Los ARNip diseñados a medida se han utilizado para deshabilitar la expresión de genes específicos con el fin de estudiar su función in vivo e in vitro. Tanto los ARNip como los miARN están siendo investigados como posibles herramientas terapéuticas para interferir con los ARN cuya expresión conduce al cáncer u otras enfermedades.
234 ARNIp Regulación Post Transcripcional
235 ¿El ARNIp Cooptó la estrategia de RISC para Basurar ARN Corrupto
Para un ejemplo echa un vistazo a un video de Youtube de resultados inesperados de un experimento de ARNi en este enlace. En el experimento descrito, se utilizó ARNi para bloquear la expresión embrionaria del gen del ortodentículo (odt) que normalmente se requiere para el crecimiento de cuernos en un escarabajo de estiércol. El efecto de esta mutación knock-out fue, como se esperaba, para prevenir el crecimiento del cuerno. Sin embargo, lo inesperado fue el desarrollo de un ojo en medio de la cabeza del escarabajo ('tercer ojo' en la micrografía).
El 3er ojo no sólo parece un ojo, sino que es funcional. Esto se demostró previniendo el desarrollo normal del ojo en mutantes deficientes odt. El 3er ojo apareció..., ¡y respondía a la luz! Ten en cuenta que este era un escarabajo con un 3er ojo, ¡no Drosophila! Para citar a Justin Kumar, de la Universidad de Indiana, quien aunque no participó en la investigación, afirmó que “... las lecciones aprendidas de Drosophila pueden no ser tan aplicables en general como yo u otros Drosófilistas, me gustaría creer... La capacidad de usar ARNi en sistemas modelo no tradicionales es un gran avance eso probablemente conducirá a una visión más equilibrada del desarrollo”.
2. Micro ARN (miARN)
Los miARN se dirigen a los ARN celulares endógenos no deseados para su degradación. Se transcriben a partir de genes que ahora se sabe que están ampliamente distribuidos en eucariotas. La ruta desde la transcripción del pre-miARN a través del procesamiento y la degradación del ARNm diana se ilustra en la página siguiente.
A medida que se transcriben, los pre-miARN se pliegan en una estructura tallo-bucle que se pierde durante el procesamiento citoplásmico. Al igual que los siARN, los miARN maduros se combinan con proteínas RISC. El complejo proteína RISC-miARN se dirige a ARNm o ARNm viejos o que ya no son necesarios dañados durante la transcripción.
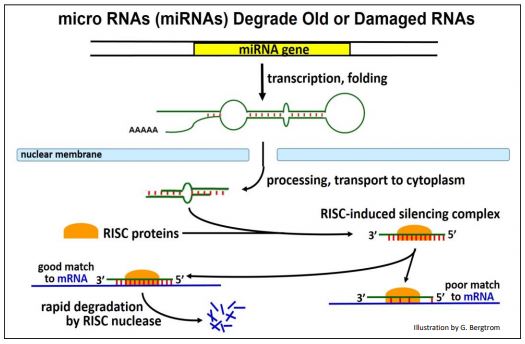
Se estima que 250 miARN en humanos pueden ser suficientes para unirse H a diversos ARN diana; solo se degradarán dianas con fuerte complementariedad con un complejo de proteína RISC-miARN.
D. ARN largos no codificantes
Los ARN largos no codificantes (lncRNAs) son otra clase más de ARN eucariotas. Incluyen transcritos de ADN antisentido, intrónico, intergénico, pseudogénico y retroposón. Los retroposones son un tipo de transposón, o elemento de ADN móvil; los pseudogenes son genes reconocibles con mutaciones que los hacen no funcionales. Si bien algunos lncRNAs pueden llegar a ser transcriptos incidentales que la célula simplemente destruye, otros tienen un papel en la regulación de la expresión génica.
Un LncRNA recientemente descubierto es Xistar que, junto con el producto del gen Xist, se requiere para formar cuerpos Barr. Los cuerpos Barr se forman en hembras humanas cuando se inactiva uno de los cromosomas X en las células somáticas. Para una revisión de los LNCRNAs, véase Lee, J.T. (2012. Regulación Epigenética por ARN Largos No Codificantes; Science 338, 1435-1439).
Un artículo aún más reciente (en LncRNAs y SMORF) resume el descubrimiento de que algunos ARN largos no codificantes contienen marcos de lectura abiertos cortos (SMORF) que en realidad se traducen en péptidos cortos de más de 30 aminoácidos. ¿Quién sabe? ¡De hecho, el genoma humano puede contener más de 21,000-25,000 genes que codifican proteínas!
E. ARN circulares (circRNA)
Aunque se descubrieron hace más de 20 años, los ARN circulares (circRNAs) se elaboran en diferentes tipos de células eucariotas. Haga clic en RNAs circulares (CircRNA) para conocer más sobre este peculiar resultado del empalme alternativo. Al principio los circRNAs fueron difíciles de aislar. Cuando se aislaron, los circRNAs contenían secuencias exónicas “codificadas” y, por lo tanto, se pensó que eran errores no funcionales del corte y empalme de ARNm.
De hecho, los circRNAs son bastante estables. Sus niveles pueden subir y bajar en patrones que sugieren que son moléculas funcionales. Los niveles de un circRNA, llamado CirCrims1, aumentan específicamente durante el desarrollo neural. En los ratones, otros circRNAs se acumulan durante la formación de sinapsis, probablemente influyendo en cómo estas neuronas finalmente se desarrollarán y funcionarán. Así, los circRNAs no parecen ser 'errores moleculares'. De hecho, ¡los errores en su propia síntesis pueden estar correlacionados con la enfermedad! La especulación sobre las funciones de los circRNAs también incluye papeles en la regulación génica, particularmente los genes o ARNm de los que ellos mismos se derivan.
F. “ADN basura” en perspectiva
No hace mucho, pensábamos que menos del 5% de un genoma eucariota estaba transcrito (es decir, en ARNm, ARNr y ARNt), y que gran parte del genoma no transcrito cumplía una función estructural... o ninguna función en absoluto. Este último, etiquetado como ADN basura, incluía secuencias intergénicas indescriptas, pseudogenes, transposones 'muertos', largos tramos de ADN intrónico, etc. Así, el ADN basura era ADN del que podíamos prescindir. Se pensaba que los ADN basura eran jinetes accidentales en nuestros genomas, los autoestopistas recogidos en el camino evolutivo.
Mientras que los genes de miARN son una pequeña proporción de un genoma eucariota, su descubrimiento y el de los ARN lnc más abundantes sugieren una cantidad mucho mayor de ADN funcional en el genoma. ¿Podría haber de hecho, no tal cosa como “ADN basura”? Continúa el debate sobre cuánto de nuestro ADN genómico es una reliquia de experimentos evolutivos pasados y sin propósito genético. Lee todo al respecto en Junk DNA - no tan inútil después de todo y Solo 8.2% del ADN humano es funcional.
Quizás necesitamos repensar lo que significa que el ADN sea “basura” o que esté sin “propósito genético”. El mantenimiento de más del 90% de nuestro propio ADN sin ningún propósito genético conocido seguramente tiene un costo energético. Al mismo tiempo, todo ese ADN es esencial para la selección futura, fuente de la diversidad requerida para la supervivencia a largo plazo. La misma selección natural que recoge secuencias de ADN de 'autoestopista', como hemos visto, puede en algún momento, ponerlas a trabajar!
G. El metiloma de ARN
¡Llama a esto epi-transcriptoma de ARN si quieres! Recordemos que los grupos metilo dirigen la escisión de los ARN ribosómicos a partir de transcritos eucariotas de pre-ARN 45S, entre otros transcritos, también se modifican postranscripcionalmente. Conocidas desde la década de 1970, se pensó que tales modificaciones no eran funcionales. Pero, ¿son ellos?