7.7: Telómeros
- Page ID
- 52987
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Si existe un mecanismo para reconocer los cabos sueltos del ADN, ¿qué pasa con los extremos de cada cromosoma eucariota? Son cromosomas lineales, entonces tienen extremos, ¿verdad? ¿Qué impide que los sistemas de reparación de doble hebra los reconozcan mal como ADN roto y concatenen todos los cromosomas juntos? Curiosamente, la respuesta a esta pregunta está íntimamente ligada a la respuesta al problema de la replicación final, al que se aludió muy brevemente en nuestra descripción de la replicación.
El problema de replicación final es uno que afecta a todos los cromosomas lineales. Se reduce a un simple hecho: se necesita un cebador de ARN para iniciar cualquier replicación del ADN. Entonces en el extremo 5' de cada cadena hay un cebador de ARN (Figura\(\PageIndex{24}\) en amarillo) que se elimina por el proceso de corrección de errores. Así, con cada ronda de replicación, se pierde información del extremo 5' de cada hebra de cada cromosoma.
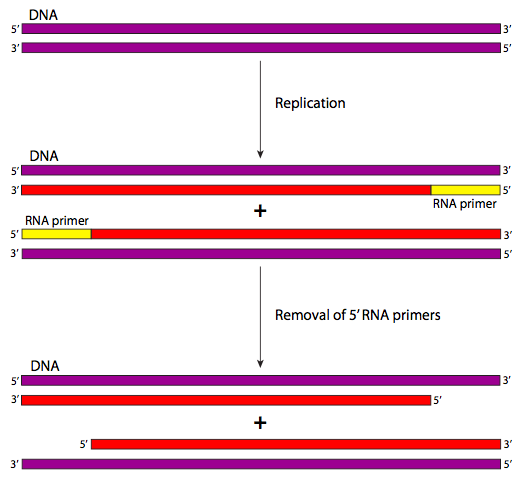
Eventualmente, se pierden genes cruciales y la célula morirá; lo más probable es que muchas funciones celulares se vean comprometidas mucho antes de que eso suceda. La solución al problema de replicación final podría considerarse más un tratamiento de los síntomas que una cura, para usar una analogía con la medicina. En definitiva, durante las etapas muy tempranas de la vida de un organismo, se agrega mucho ADN no codificante en los extremos del ADN para que a medida que la célula y su progenie continúan reproduciéndose, los nucleótidos no afectan a ningún gen funcional. Este proceso es catalizado por la enzima telomerasa.
La telomerasa es una holoenzima grande que actúa como transcriptasa inversa, leyendo un molde de ARN autónomo para agregar la secuencia del telómero a los extremos 3' de los cromosomas lineales. Tenga en cuenta que esto no agrega a los extremos 5' - como se mencionó anteriormente, no hay nada que hacer sobre los extremos 5' directamente. Sin embargo, mientras las telomerasas están activas, los extremos 3' pueden extenderse, y así cuando se elimina el cebador del extremo 5', la secuencia perdida (para siempre) de esa cadena de ADN es solo una repetición telomérica y no algo más útil. Las repeticiones están bien conservadas a través de eucariotas, y casi completamente conservadas en todas las especies de mamíferos (se muestra en la Figura\(\PageIndex{25}\)).
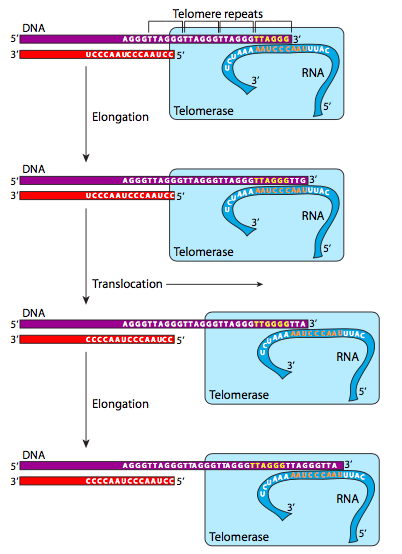
En metazoos, la actividad de la telomerasa es alta en las etapas embrionarias de la vida, pero es prácticamente inexistente en adultos excepto en los tipos de células que deben proliferar constantemente (por ejemplo, células sanguíneas y epiteliales). La actividad de la telomerasa está regulada principalmente por la expresión del gen TERT (transcriptasa inversa de telomerasa) aunque la construcción de la telomerasa completa también requiere la expresión del gen TERC (el ARN de la telomerasa, también abreviado TR) y la disquerina. En términos generales, el número de repeticiones teloméricas que se colocan en un cromosoma en el desarrollo temprano determina el número de repeticiones de ADN y divisiones celulares que puede sufrir la célula antes de sucumbir a la apoptosis (muerte celular programada). Los experimentos en células en cultivo demuestran una fuerte correlación entre la longitud de los telómeros y la longevidad, y se sabe que las células tomadas de personas con la enfermedad de envejecimiento prematuro, progeria, tienen telómeros relativamente cortos.
Por el contrario, las células cancerosas casi universalmente tienen expresión no regulada de telomerasa. Dado que una característica definitoria de las células cancerosas es la capacidad de proliferar rápida e indefinidamente, volver a encender la telomerasa es, no es sorprendente, un aspecto importante de la carcinogénesis. Por lo tanto, es una diana para los tratamientos anticancerosos; sin embargo, hasta la fecha ninguna terapia dirigida a telomerasa ha demostrado ser efectiva.
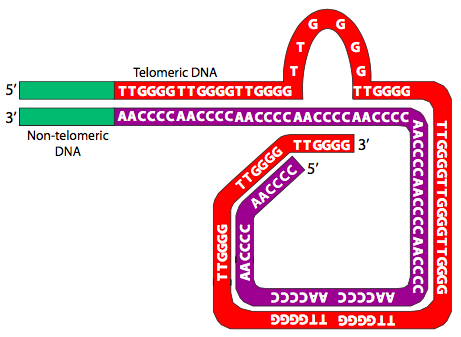
Ahora que conocemos los telómeros, la pregunta que inició esta sección se vuelve aún más problemática: con estos voladizos de secuencia repetidos, ¿cómo se impide que los cromosomas se conecten de extremo a extremo a través de un proceso de reparación de doble hebra? En parte debido a sus secuencias repetidas, los telómeros son capaces de formar casquetes terminales y proteger los extremos cromosómicos. Los telómeros protegen los extremos de cada cromosoma uniéndose a proteínas protectoras y formando estructuras complejas. Las proteínas de unión al extremo del telómero (TEBP) se unen al extremo 3' que sobresale del telómero. Otras proteínas de protección, como la TRF1 y TRF2 de mamíferos (factores de unión a repetición de telómeros) no solo se unen al telómero, sino que ayudan a organizarlo en grandes estructuras en bucle conocidas como bucles T (Figura\(\PageIndex{26}\)).
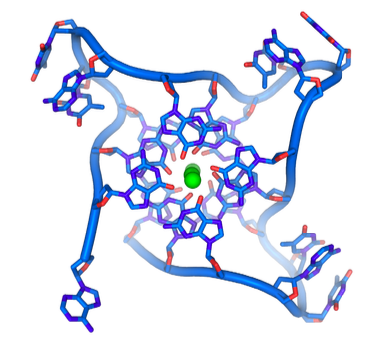
Finalmente, los extremos del lazo T se estabilizan adicionalmente mediante la formación de cuartetos G (Figura\(\PageIndex{27}\)). Los cuartetos G son tetrámeros cíclicos que se pueden formar en secuencias con cuatro residuos de guanina consecutivos, los cuales se unen de hidrógeno entre sí para formar una forma cuadrada enlazada estabilizada por un ion metálico en el centro. Además, en casos como el telómero, en los que se repiten tales secuencias, los cuartetos G pueden apilarse y asociarse tridimensionalmente, aumentando su estabilidad.