12.6: Transporte en el Citoesqueleto
- Page ID
- 53358

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
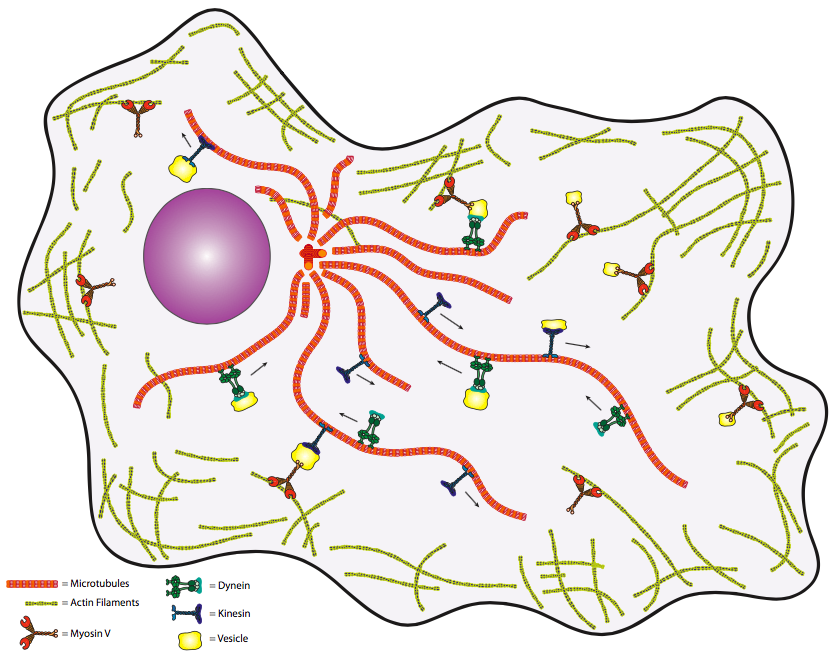
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Si bien puede ser útil pensar en estas estructuras citoesqueléticas como análogas a un esqueleto animal, quizás una mejor manera de recordar la ubicación relativa de los microtúbulos y microfilamentos es por su función en el transporte de carga intracelular de una parte de la célula a otra. Por esa analogía, podríamos considerar los microtúbulos como un sistema de vías férreas, mientras que los microfilamentos se parecen más a las calles. Por la misma analogía, podemos sugerir que la red de microtúbulos y la red de microfilamentos están conectadas en ciertos puntos para que cuando la carga llegue a su destino general por microtúbulos (ferrocarril), entonces pueda ser llevada a su dirección específica por microfilamento. Extendamos un poco más esta analogía. Si los microtúbulos y microfilamentos son las vías y las calles, entonces ¿cuáles son los trenes y camiones? Ah, una pregunta astua, Saltamontes. En los microtúbulos, los “trenes” son una de las dos familias de motores moleculares: las quinesinas y las dineínas.
Podemos generalizar un poco y decir que las quinesinas conducen hacia el extremo (+) (hacia la periferia de la célula) mientras que las dineínas van hacia el extremo (-) (hacia el MTOC). En los microfilamentos de actina, los motores moleculares son proteínas de la familia de la miosina. En este punto, terminan las analogías, ya que el funcionamiento de estos motores moleculares es muy diferente al de la locomoción en tren o camión. Por último, se podría cuestionar la necesidad biológica de tal sistema de transporte. Nuevamente, si nos analogicamos con el transporte humano, entonces podríamos decir que el transporte vía difusión simple es similar a personas que llevan paquetes al azar alrededor de la celda. Es decir, las entregas eventualmente se harán, pero no querrás contar con este método para materiales críticos de tiempo. Por lo tanto, se necesita un sistema dirigido de alta velocidad para mantener vivas las células (particularmente las células eucariotas más grandes).
Aunque este tipo de transporte ocurre en todas las células eucariotas, un caso particularmente bien estudiado es el transporte axonal (también llamado transporte axoplásmico) en neuronas. Aquí, el transporte de materiales desde el cuerpo celular (soma) hasta las puntas de los axones a veces puede atravesar distancias muy largas hasta varios metros en animales más grandes, y debe hacerlo de manera oportuna. El transporte axonal generalmente se clasifica como anterógrado (desde soma hasta axón terminal) o retrógrado (desde terminales posteriores). Los tipos de material transportado en estas dos direcciones son muy diferentes: gran parte del transporte anterógrado son bloques de construcción de proteínas para extender el axón o vesículas sinápticas que contienen neurotransmisores; el transporte retrógrado es principalmente vesículas endocíticas y moléculas de señalización. El transporte axonal también se clasifica como rápido y lento. El transporte lento es principalmente el movimiento de proteínas directamente unidas a los motores, y pueden pasar de 100 mm por día (SCa, componente lento a) hasta 3 mm/día (scB). En comparación, el transporte rápido es generalmente movimiento de vesículas, y puede variar de 50 a 400 mm/día. El mecanismo de transporte lento se había debatido desde hace más de una década hasta el año 2000, cuando la visualización directa de neurofilamentos marcados fluorescentemente en el transporte mostró que el movimiento real de las proteínas era muy similar al movimiento en el transporte axonal rápido, pero hubo muchas pausas en el transporte, un” stop and go” en lugar de moverse de origen a destino continuamente.
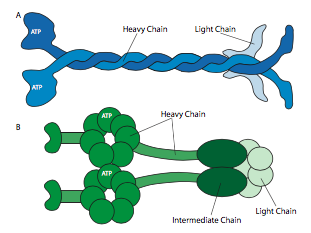
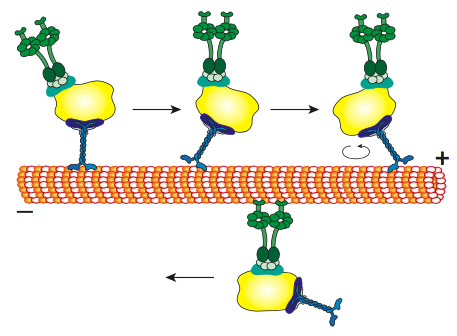
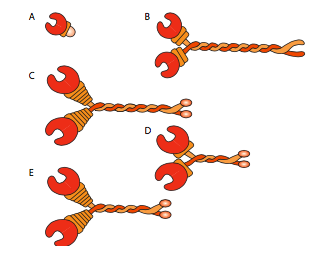
Todas las kinesinas y dineínas tienen algunos puntos en común clave. Hay una “cabeza” catalítica liberadora de energía conectada a una región de bisagra o cuello que permite que la molécula se flexione o “escalone”, y hay una cola portadora de carga más allá de eso (Figura\(\PageIndex{8}\)). La cabeza de una cinesina o dineína cataliza la hidrólisis del ATP, liberando energía para cambiar su conformación en relación con el cuello y la cola de la molécula, lo que le permite liberar temporalmente su agarre sobre el microtúbulo, girar sus “caderas” alrededor para plantarse a un “paso” de distancia, y volver a unirse al microtúbulo (Figura \(\PageIndex{9}\)). En los microfilamentos de actina, las miosinas, de las cuales también hay muchos tipos (algunas representadas en la Figura\(\PageIndex{10}\)) son los motores moleculares. Su movimiento es diferente al de las dineinas y quinesinas, como se describirá en la siguiente sección, pero también utiliza la energía de la hidrólisis de ATP para proporcionar energía para los cambios conformacionales necesarios para el movimiento. Hemos introducido los motores, pero considerando la enorme diversidad en las moléculas que necesitan ser transportadas alrededor de una celda, sería imposible que los motores se unan directamente a todos ellos. De hecho, los motores se unen a su carga a través de moléculas adaptadoras que unen el motor por un lado, y una molécula de carga o vesícula en el otro. En el capítulo de transporte vesicular se discutió un mayor examen de la carga y el enrutamiento de la carga por marcadores de dirección (SNares).