
14.1: Desarrollo Embrionario
- Page ID
- 57037

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
En los animales, generalmente se pueden distinguir 4 etapas de desarrollo embrionario.
- Escote
- Patrones
- Diferenciación
- Crecimiento
Escote
La mitosis y citocinesis del cigoto, una célula inusualmente grande, produce un número creciente de células más pequeñas, cada una con una copia exacta del genoma presente en el cigoto. Sin embargo, los genes del cigoto no se expresan al principio. Las actividades tempranas de escisión están controladas por el genoma de la madre; es decir, por los ARNm y las proteínas que depositó en el óvulo no fertilizado. En los humanos, el cambio ocurre después de que se hayan producido 4-8 células; en ranas no hasta que se hayan producido miles de células. La escisión termina con la formación de una blástula.
Patrones
Durante esta fase, las células producidas por escisión se organizan en capas y masas, un proceso llamado gastrulación. El patrón del futuro animal aparece:
- de delante a atrás (el eje anterior-posterior)
- lado posterior y lado del vientre (su eje dorsal-ventral)
- lados izquierdo y derecho.
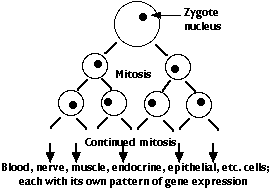
El genoma del cigoto contiene todos los genes necesarios para elaborar los cientos de diferentes tipos de células que conformarán al animal completo. Hay dos categorías principales de estos genes:
- Genes “domésticos” = aquellos que codifican los ARN y proteínas que necesitan todo tipo de células. Ejemplos:
- genes para ARNt, ARNr
- genes que codifican las enzimas de la glucólisis.
- genes específicos de tejido = aquellos que codifican ARNm y por lo tanto proteínas que son utilizadas por uno o unos pocos tipos específicos de células. Ejemplos:
- genes para la hemoglobina expresada en los precursores de los glóbulos rojos
- el gen de la insulina expresado en las células beta de los islotes de Langerhans
Sin embargo, cada célula descendiente del cigoto ha sido producida por mitosis y así contiene el genoma completo del organismo (con muy pocas excepciones).
Dos pruebas:
- Dolly - Dolly es la oveja que se formó insertando un núcleo de una sola célula de una oveja adulta en un huevo de oveja enucleada. Ella prueba que la célula del adulto no había perdido ninguno de los genes necesarios para construir todos los tejidos de una oveja.
- Experimentos de atado de huevos de Spemann - Muchos años antes, el embriólogo alemán Hans Spemann demostró la misma verdad. Usó mechones de pelo de bebé para atar bucles alrededor de huevos de salamandra fertilizados. Aunque la mitad del huevo con el núcleo comenzó a escindirse normalmente, el otro lado no comenzó a escindirse hasta que finalmente un núcleo se deslizó por el nudo. Siempre y cuando el huevo estuviera atado para que ambas mitades contuvieran algo de la media luna gris, la segunda mitad comenzó la escisión normal y finalmente produjo un segundo renacuajo (derecha). Incluso después de 5 divisiones mitóticas del núcleo del cigoto (la etapa de 32 células), el genoma completo todavía estaba disponible en cada núcleo descendiente.


- Un óvulo fertilizado es mucho más grande que las células normales del cuerpo de un animal. Algunos (por ejemplo, un huevo de gallina) son realmente enormes. El huevo de rana tiene un volumen 1.6 millones de veces más grande que una célula de rana típica. La foto es de un embrión de rana de 16 células. Esta masa de células no es mayor que el huevo original. Los huevos de los mamíferos son más pequeños, pero incluso son más grandes de lo que serán sus células descendientes.
- El citoplasma del óvulo fertilizado no es homogéneo. Contiene gradientes de ARNm y proteínas. Estos son los productos de los genes de la madre y fueron depositados en el óvulo por ella.
- La escisión del óvulo fertilizado lo divide en miles de células de tamaño normal. Cada uno contiene un núcleo descendiente del núcleo cigoto.
- Pero cada núcleo se encuentra dividido en citoplasma que contiene una mezcla particular de ARNm y proteínas.
- Cuando la blástula de rana ha producido unas 4 mil células, comienza la transcripción y traducción de sus genes nucleares (y se destruyen las moléculas de ARNm de la madre, que hasta ahora han sido la fuente de toda la síntesis de proteínas).
- Los genes que son expresados por el núcleo en una célula dada están regulados por las moléculas, en su mayoría factores de transcripción proteica y microARN (miRNAs), que se encuentran en el citoplasma que rodea ese núcleo.
- Una vez que se lanza un patrón específico de célula de expresión génica, esa célula puede liberar moléculas que regulan los genes de las células cercanas.
- De esta manera, se sientan las bases para la construcción de un organismo con cientos de tipos de células diferenciadas —cada una en su correcta ubicación y realizando sus funciones correctas.
Xenopus

- Durante la formación del huevo, las moléculas de ARNm que codifican la proteína VEGT se depositan en el polo vegetal de la célula.
- Las células que allí se forman durante la escisión traducen el ARNm a la proteína VEGT.
- VEGT es un factor de transcripción que activa genes que producen miembros de la familia del factor de crecimiento transformante beta (TGF-β) (por ejemplo, activina).
- Estas proteínas son necesarias para que las células comiencen por el camino de convertirse en mesodermo.
- Algunas de esas celdas, a su vez, se convertirán en el organizador de Spemann.
- Posteriormente, el organizador Spemann segregará moléculas que inducen a las células ectodérmicas por encima de ellas a desarrollarse en los tejidos del cerebro y la médula espinal.
Demostración
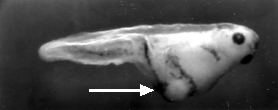

Inyectar la parte anterior del óvulo fertilizado con mRNA de nanos. El resultado: otra larva doble posterior.
Hacer moscas hembra de la fruta que sean transgénicas para un gen recombinante que contenga:
- el gen para los nanos
- acoplado a la señal 3' de dirección anterior del gen bicoide.
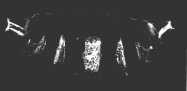

A la derecha se muestra una larva normal. El objeto brillante en el extremo derecho de la larva normal y en ambos extremos de la larva posterior doble es la punta de la cola. Estas micrografías son cortesía de Elizabeth Gavis y Ruth Lehmann, en cuyo laboratorio se realizó la tercera demostración.
El Caracol de Barro
El caracol de barro, Ilyanassa obsoleta, es un pequeño gasterópodo que vive en llanuras de barro a lo largo de la costa atlántica
Al igual que otros protostomos, la escisión del cigoto produce células hijas que ya están comprometidas con su destino. En otras palabras, incluso ya en la etapa de dos células, las células ya no son totipotentes. A diferencia de los humanos y otros deuterostomas, entonces, no se pueden formar gemelos idénticos.
En el número 12 de diciembre de 2002 de Nature, J. David Lambert y Lisa Nagy reportaron otro mecanismo por el cual dos células hijas se comprometen con destinos diferentes a pesar de que han heredado el mismo genoma.
Rastrearon la distribución en las células de embriones tempranos de los ARN mensajeros (ARNm) que codifican 3 proteínas que se sabe que son importantes en el desarrollo de otros animales como Xenopus y Drosophila.
- IoEve, que es la versión de I lyanassa o bsoleta de par saltado (eve) en Drosophila;
- IoDPP, que es la versión del caracol de
- decapentapléjico (dpp) en Drosophila y los genes que codifican
- b una m p roteínas orfogénicas (BMP2 y BMP4) en vertebrados
- IoTLD, que codifica la versión del caracol de una proteína llamada tolloide en Drosophila.
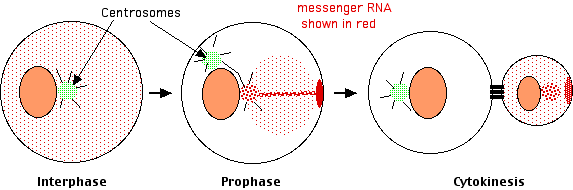
Lambert y Nagy encontraron que
- en la interfase los ARN mensajeros se distribuyeron difusamente por todo el citosol, pero
- a medida que la célula se preparaba para la escisión, los ARNm se recolectaron solo en uno de los ahora pares de centrosomas. Allí se recolectaron viajando a lo largo de los microtúbulos que irradian desde el centrosoma.
- A medida que continuaba la escisión, los ARNm se movían del centrosoma a una mancha en la superficie interna de la membrana plasmática. Llegaron allí viajando a lo largo de filamentos de actina.
- En la citocinesis, este parche de ARNm acumulados se incorporó exclusivamente a la célula hija más pequeña.
La clasificación del centrosoma (de proteínas en este caso) también juega un papel en la determinación de si las células embrionarias de Caenorhabditis elegans permanecen en la línea germinal o se convierten en las células somáticas del gusano.
¿Qué viene después?
El desarrollo en Xenopus y Drosophila pasa a través de tres fases bastante diferentes (aunque a menudo superpuestas):
- Establecer los ejes principales (anterior-posterior; dorsal-ventral; izquierda-derecha). Esto se hace por gradientes de ARNm y proteínas codificadas por los genes de la madre y colocadas en el óvulo por ella. Se ha discutido aquí.
- Establecer las partes principales del cuerpo como la notocorda y el sistema nervioso central en vertebrados y los segmentos en drosófilaEstos están dirigidos por genes del propio cigoto.
- Relleno de los detalles; es decir, construir los diversos órganos del animal.