
15.4M: Presentación de antígeno
- Page ID
- 56858

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Los antígenos son macromoléculas que inducen una respuesta inmune en el organismo. Los antígenos pueden ser proteínas, polisacáridos, conjugados de lípidos con proteínas (lipoproteínas) y polisacáridos (glicolípidos). La mayor parte de esta página describirá cómo se presentan los antígenos proteicos al sistema inmunitario. Al final se mencionará la presentación de antígenos lipídicos y polisacáridos. Será útil distinguir entre dos casos limitantes.
Antígenos que ingresan al cuerpo desde el ambiente; estos incluirían macromoléculas inhaladas (por ejemplo, proteínas en pelos de gato que pueden desencadenar un ataque de asma en personas susceptibles), macromoléculas ingeridas (por ejemplo, proteínas de mariscos que desencadenan una respuesta alérgica en personas susceptibles) y moléculas que son introducido debajo de la piel (por ejemplo, en una astilla o en una vacuna inyectada). Alternativamente, se pueden generar antígenos dentro de las células del cuerpo; estos incluirían proteínas codificadas por los genes de virus que han infectado una célula y proteínas aberrantes que son codificadas por genes mutantes; tales como genes mutados en células cancerosas. En todos los casos, sin embargo, la respuesta inmune inicial a cualquier antígeno requiere absolutamente que el antígeno sea reconocido por un linfocito T (“célula T”). La verdad de esta regla está claramente demostrada en el SIDA: las infecciones (virales o fúngicas o bacterianas) que tantas veces reclaman la vida de los pacientes con SIDA lo hacen cuando el paciente ha perdido prácticamente la totalidad de sus células T CD4+. Las dos categorías de antígenos son procesadas y presentadas a las células T por mecanismos muy diferentes.
Antígenos exógenos
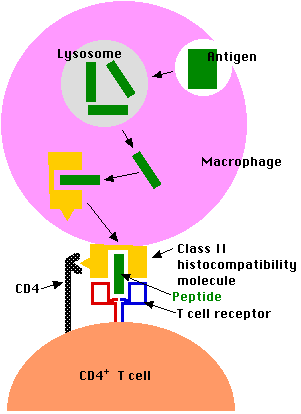
Los antígenos exógenos (inhalados, ingeridos o inyectados) son captados por las células presentadoras de antígeno (APC). Estas incluyen células fagocíticas como células dendríticas y macrófagos y linfocitos B (“células B”) que son responsables de producir anticuerpos contra el antígeno. Células presentadoras de antígeno
- engullir el antígeno por endocitosis
- el endosoma se fusiona con un lisosoma donde el antígeno se degrada en fragmentos (por ejemplo, péptidos cortos)
- estos péptidos antigénicos se muestran entonces en la superficie de la célula enclavada dentro de una molécula de histocompatibilidad de clase II.
- pueden ser reconocidos por células T CD4 +
El Camino Clase I
Las moléculas de histocompatibilidad Clase I son proteínas transmembrana expresadas en la superficie celular. Como todas las proteínas transmembrana, son sintetizadas por ribosomas en el retículo endoplásmico rugoso (RER) y ensambladas dentro de su lumen. Hay tres subunidades en cada molécula de histocompatibilidad de clase I:
- el polipéptido transmembrana (llamado la “cadena pesada”)
- el péptido antigénico
- beta-2 microglobulina
Todos estos deben estar presentes dentro de la luz del retículo endoplásmico para que se ensamblen correctamente y se muevan a través del aparato de Golgi hasta la superficie celular. El problema: las proteínas codificadas por los genes de un virus infeccioso se sintetizan en el citosol. ¿Cómo introducirlos en el retículo endoplásmico?
La Solución: TAP (= t ransporter a ssociated con antígeno p rocessing).
- Las proteínas virales en el citosol son degradadas por los proteasomas en péptidos virales.
- Los péptidos son captados por proteínas TAP incrustadas en la membrana del retículo endoplásmico.
- Usando la energía del ATP, los péptidos son bombeados al lumen del retículo endoplásmico donde se ensamblan con el polipéptido transmembrana y beta-2 microglobulina.
- Este complejo trimolecular luego se mueve a través del aparato de Golgi y se inserta en la membrana plasmática.
- El complejo puede unirse por una célula T con un receptor (TCR) capaz de unirse al péptido y porciones flanqueantes de la molécula de histocompatibilidad (el hot dog en el bollo) y moléculas CD8 que se unen al receptor CD8 (mostrado anteriormente como un hemisferio gris) en el molécula de histocompatibilidad.
El Camino Clase II
Las moléculas de histocompatibilidad Clase II consisten en dos polipéptidos transmembrana y una tercera molécula enclavada en el surco que forman. Los tres componentes de este complejo deben estar presentes en el retículo endoplásmico para su correcto ensamblaje. Pero los péptidos antigénicos no son transportados al retículo endoplásmico, por lo que una proteína llamada cadena invariante (” Ii “) ocupa temporalmente el surco.
Los pasos:
- Las dos cadenas de la molécula de clase II se insertan en la membrana del retículo endoplásmico.
- Se unen (en su surco) a una molécula de cadena invariante.
- Este complejo trimolecular es transportado a través del aparato de Golgi y a vesículas llamadas lisosomas.
Mientras tanto, el material antigénico extraño es envuelto por endocitosis formando endosomas. Estos también se fusionan con lisosomas. Entonces,
- El antígeno se digiere en fragmentos.
- Se digiere la cadena invariante (Ii).
- Esto libera el surco para la ocupación por el fragmento antigénico.
- Las vesículas se desplazan a la membrana plasmática y el complejo se muestra en la superficie celular.
- El complejo puede estar unido por una célula T con
- un receptor (TCR) capaz de unirse al péptido y porciones flanqueantes de la molécula de histocompatibilidad (el hot dog en el bollo) y
- Las moléculas CD4 que se unen al receptor CD4 (mostrado arriba como un triángulo amarillo) se encuentran en todas las moléculas de histocompatibilidad de clase II.
Interconexiones entre las vías Clase I y Clase II
Presentación Cruzada: Transferencia de Antígenos Exógenos a la Vía Clase I
La presentación cruzada es la transferencia de antígenos extracelulares como bacterias, algunos antígenos tumorales y antígenos en células infectadas por virus a la vía de clase I para la estimulación de células T citotóxicas (CTL) CD8 +. Solo ciertas células presentadoras de antígeno (APC) “profesionales” como las células dendríticas pueden hacer esto: usan las vías de presentación de antígeno de clase I y de clase II.
La presentación cruzada después de la infección por virus es importante porque:
- La mayoría de los virus infectan células distintas de las APC (por ejemplo, células hepáticas, células epiteliales del pulmón) (y, por supuesto, son intracelulares en estas).
- Si bien los antígenos virales que se muestran en la superficie de cualquier célula infectada pueden servir como dianas para las células T citotóxicas (CTL),
- la falta de moléculas coestimuladoras en la superficie celular las convierte en pobres estimulantes para el desarrollo de clones de CTL en primer lugar.
Sin embargo, cuando una célula infectada muere, puede ser engullida por un APC profesional, y los antígenos dentro de ella pueden ingresar a la vía de clase I. Un mecanismo:
- La célula muerta es engullida por endocitosis.
- El endosoma que forma se fusiona con un lisosoma y comienza la degradación de la célula muerta.
- Los antígenos pasan al citosol y se degradan en proteasomas.
- Los péptidos formados son luego recogidos por TAP y se insertan en moléculas MHC clase I y se muestran en la superficie celular, junto con las moléculas coestimuladoras necesarias para iniciar una expansión clonal vigorosa de células T citotóxicas CD8 +.
Desviación de Antígenos de la Vía Clase I a la Clase II
La autofagia proporciona un mecanismo mediante el cual las células pueden transferir antígenos endógenos (intracelulares) a la vía de clase II, por ejemplo
- autoproteínas para poder eliminar linfocitos T CD4+ con receptores capaces de atacarlos y así potencialmente capaces de causar autoinmunidad
- proteínas sintetizadas por un virus infeccioso. De esta manera, la infección viral puede generar células T CD4+ así como células T citotóxicas (CD8 +)
Linfocitos B: Un Caso Especial

Los linfocitos B son tanto células receptoras de antígeno como presentadoras de antígeno. Se unen a antígenos intactos (por ejemplo, partículas de virus, proteínas) con su receptor de células B (BCR). Pueden entrar en contacto con estos antígenos encontrándolos en la linfa circundante o al ser presentados por macrófagos o células dendríticas. Los linfocitos B procesan el antígeno por la vía de clase II para su presentación a células T.
El proceso:
- Las células B engullan antígeno por endocitosis mediada por receptores
- Los receptores de células B para antígeno (BCR) son anticuerpos anclados en la membrana plasmática.
- La afinidad de estos por un epítopo sobre un antígeno puede ser tan alta que la célula B puede unirse e internalizar el antígeno cuando está presente en fluidos corporales en concentraciones miles de veces más pequeñas de las que necesitaría un macrófago.
- Las etapas restantes del procesamiento del antígeno ocurren por la misma vía de clase II descrita anteriormente para los macrófagos productores
- fragmentos de antígeno mostrados en la superficie celular enclavada en el surco de las moléculas de histocompatibilidad de clase II.
- Una célula T CD4+ que reconoce el antígeno mostrado es estimulada para liberar linfocinas.
- Estos, a su vez, estimulan a la célula B para que entre en el ciclo celular.
- Debido al papel que desempeñan en la estimulación de las células B, estas células T CD4+ se denominan células T auxiliares (” Th “).
- La célula B se convierte en un clon de células (llamadas células plasmáticas)
- Estos sintetizan receptores (BCR) con el sitio de unión idéntico para el epítopo pero sin la cola transmembrana.
- Estos anticuerpos son secretados a los alrededores.
Antígenos lípidos y polisacáridos
Antígenos Lípidos
- Los antígenos lipídicos se presentan a las células T mediante moléculas de superficie celular designadas CD1 (“agrupación de diferenciación” 1).
- Las células presentadoras de antígeno expresan varias formas diferentes de CD1 en su superficie. Cada uno está probablemente especializado para unirse a un tipo particular de antígeno lipídico (por ejemplo, lipopéptido vs glicolípido).
- La superficie expuesta de las moléculas CD1 forma un surco de unión al antígeno muy parecido al de las moléculas del MHC, excepto que
- los aminoácidos en el surco son más hidrófobos que los de las moléculas del MHC.
- Al igual que los antígenos proteicos, los antígenos lipídicos también se presentan como fragmentos, es decir, como un “perrito caliente en un bollo”.
Antígenos polisacáridos
Algunos polisacáridos bacterianos ingeridos por APC
- pueden degradarse en sus lisosomas
- y presentado a células T por moléculas MHC clase II.

La unión de una célula T a una célula presentadora de antígeno (APC) por sí misma no es suficiente para activar la célula T y convertirla en una célula efectora: una capaz de, por ejemplo,
- matar el APC (CD8 + linfocitos T citotóxicos [CTL])
- llevar a cabo reacciones inmunes mediadas por células (células CD4 + Th1)
- proporcionar ayuda a las células B (células CD4 + Th2)
Para activarse, la célula T no solo debe unirse al epítopo (MHC-péptido) con su TCR sino también recibir una segunda señal de la APC. La recepción de esta segunda señal se llama coestimulación. Entre los más importantes de estos coestimuladores se encuentran las moléculas en la APC designadas B7 y su ligando en la célula T designado CD28. La unión de CD28 a B7 proporciona la segunda señal necesaria para activar la célula T.
Aunque las células T pueden encontrar autoantígenos en los tejidos corporales, no responderán a menos que reciban una segunda señal. De hecho, la unión de su TCR (“señal uno”) sin “señal dos” hace que se autodestruyan por apoptosis. La mayoría de las veces, las células que presentan los antígenos propios del cuerpo, ya sea
- no proporcionar la señal dos o
- transmitir una segunda señal aún no identificada que convierte la célula T en una célula T reguladora (Treg) que suprime las respuestas inmunitarias.
En cualquier caso, los resultados de autotolerancia.