
15.4R: El Sistema Complemento
- Page ID
- 56820

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
A veces la interacción de anticuerpos con antígeno es útil por sí misma. Por ejemplo,
- recubrir un virus o bacteria evitando así que se una a, e invada, una célula hospedadora (por ejemplo, anticuerpos antipolio)
- unión a una molécula de toxina (por ejemplo, toxina diftérica o tetánica) evitando así que la toxina entre en una célula donde realiza su trabajo sucio
Pero la mayoría de las veces, la unión de anticuerpos al antígeno no realiza ninguna función útil hasta y a menos que pueda activar un mecanismo efector. El sistema de complemento cumple varias funciones efectoras. El sistema del complemento proporciona la protección real de la respuesta mientras que la interacción de anticuerpos y antígeno proporciona la especificidad de la respuesta. Dicho de otra manera, los anticuerpos “dedo” al objetivo, el complemento lo destruye.
Características del sistema
- El sistema del complemento consiste en unas 30 proteínas que circulan en el plasma sanguíneo.
- La mayoría de estos son inactivos hasta que son escindidos por una proteasa que, a su vez, los convierte en una proteasa.
- Así, muchos componentes del sistema sirven como sustrato de un componente anterior y luego como enzima para activar un componente posterior.
- Este patrón de activación secuencial produce una cascada expansiva de actividad (que recuerda al funcionamiento del sistema de coagulación de la sangre).
El Camino Clásico
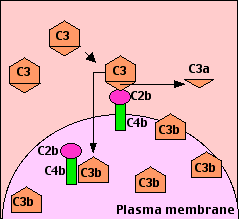
La unión del anticuerpo a su antígeno a menudo desencadena el sistema del complemento a través de la llamada vía clásica. Puede ocurrir en solución o —como se muestra aquí— cuando los anticuerpos se han unido a antígenos en una superficie celular.
Las proteínas de la vía clásica

C1
C1 existe en el suero sanguíneo como un complejo molecular que contiene:
- 6 moléculas de C1q
- 2 moléculas de C1r
- 2 moléculas de C1s
Las regiones constantes de las cadenas mu (IgM) y algunas cadenas gamma (IgG) contienen un sitio de unión para C1q. Una sola molécula de IgM es suficiente para iniciar la ruta. La IgG es mucho menos eficiente, requiriendo varias moléculas para hacerlo (6 es el óptimo, lo mismo que el número de moléculas C1q en C1).
- La unión de C1q activa C1s y C1r.
- Los C1s activados (una serina proteasa) escinden dos proteínas séricas:
- C4 se escinde en un fragmento grande
- C4b, que se une covalentemente a residuos de azúcar en glicoproteínas de la superficie celular, y un fragmento más pequeño, inactivo, de
- C4a que se difunde.
- C2 se escinde en
- C2b, que se une de forma no covalente a un sitio en C4b, dejando un fragmento más pequeño, inactivo de
- C2a que se difunde.
- El complejo de C4b•2b se llama "C3 convertasa" porque cataliza la escisión de C3. (C4b•2b también es una serina proteasa).
- C4 se escinde en un fragmento grande
C3
C3 es la proteína más abundante del sistema del complemento (~1.3 mg/ml). Por su abundancia y su capacidad de activarse, magnifica enormemente la respuesta.
- C4b•2b corta C3 en dos fragmentos principales:
- C3b, que se une covalentemente a glicoproteínas dispersas a través de la superficie celular. Los macrófagos y neutrófilos tienen receptores para C3b y pueden unirse a la célula o partícula recubierta C3B preparatoria a la fagocitosis. Este efecto califica a C3b como opsonina.
- C3a Este pequeño fragmento se libera en los fluidos circundantes. Puede unirse a receptores en basófilos y mastocitos desencadenándolos para liberar su contenido vasoactivo (por ejemplo, histamina). Debido al papel de estos materiales en la anafilaxia, C3a se llama anafilatoxina.
- Algunos de los C3b se unen a moléculas de C5 creando un cambio alostérico que los expone a la escisión por C4b•2b (que es así una “convertasa C3/C5”).
C5
La escisión de C5 por la convertasa C3/C5 inicia el ensamblaje de un conjunto de proteínas del complemento que conforman el complejo de ataque a la membrana. (El complejo de ataque a la membrana también puede estar formado por otra convertasa C5 producida por la “vía alternativa”).
El complejo de ataque de membrana
La escisión de C5 por la convertasa C3/C5 produce:
- C5a, que se libera en el entorno fluido donde
- es una potente anafilatoxina (como C3a)
- es un atrayente quimiotáctico para neutrófilos
- C5b, que sirve como anclaje para el ensamblaje de una sola molécula cada uno de C6, C7 y C8.
- El complejo resultante C5b•6•7•8 guía la polimerización de hasta 18 moléculas de C9 en un tubo insertado en la bicapa lipídica de la membrana plasmática. Este tubo forma un canal que permite el paso de iones y moléculas pequeñas. El agua ingresa a la célula por ósmosis y la célula se lisa.

La micrografía electrónica (cortesía de los doctores J. H. Humphrey y R. Dourmashkin) muestra agujeros perforados a través de la pared celular de la bacteria Gram-negativa Shigella dysenteriae por los componentes terminales del sistema del complemento. (Algunos de los agujeros son más grandes de lo esperado para los canales C9 y probablemente fueron agrandados posteriormente por la acción de la lisozima).
Funciones efectoras del complemento
La lisis celular es solo una función (y probablemente no la más importante) del sistema del complemento. El sistema de complemento actúa de varias maneras para movilizar mecanismos de defensa.
- La opsonización por C3b apunta a partículas extrañas para fagocitosis.
- La quimiotaxis por C5a atrae a las células fagocíticas al sitio de daño.
- Esto es ayudado por el aumento de la permeabilidad de los lechos capilares mediados por C3a y C5a.
- Los componentes tempranos del complemento también son importantes para solubilizar complejos antígeno-anticuerpo que ayudan en su catabolismo y eliminación del cuerpo. El fracaso de esta función puede conducir a trastornos del complejo inmune.
- Lisis de células recubiertas de anticuerpos. (En algunos casos, esto causa más daño que bien; la lisis mediada por complemento puede causar trastornos tan graves como
- Enfermedad de Rh
- anemia hemolítica inmune
- púrpura trombocitopénica inmune
- Promoviendo la formación de anticuerpos. La descomposición de C3b genera un fragmento (C3d) que se une a antígenos potenciando su captación por las células dendríticas y las células B.
(El C3d . El complejo antigénico se une al mismo receptor en las células B que usa el virus de Epstein-Barr (VEB) para ingresar a las células B, donde puede causar mononucleosis y, a veces, linfoma de Burkitt).
El Camino Alternativo
El sistema del complemento también se puede activar sin complejos antígeno-anticuerpo. Incluso en su ausencia, hay una conversión espontánea de C3 a C3b. Normalmente el C3b se inactiva rápidamente: el C3b se une a las proteínas inhibidoras y al ácido siálico presentes en la superficie de las propias células del cuerpo, y el proceso se aborta. Sin embargo, las bacterias y otros materiales extraños que pueden entrar en el cuerpo carecen de estas proteínas y tienen poco o ningún ácido siálico.

Así que el C3b
- se une a una proteína llamada Factor B formando un complejo de C3b•BB.
- C3b•bb es también una convertasa C3 que actúa sobre más C3 para formar:
- C3b•BB•C3b, que es una convertasa C5 y puede iniciar el ensamblaje del complejo de ataque de membrana.
- más C3b! Esta segunda función (mostrada aquí) crea un bucle de retroalimentación positiva, amplificando lo que podría haber comenzado como una pequeña reacción (la formación de C3b por cualquiera o ambas vías clásica y alternativa) en una producción masiva de C3b.
Regulación de la actividad del complemento
El potencial explosivo del sistema de complemento requiere que se mantenga bajo un estricto control. Se conocen al menos 12 proteínas que hacen esto. Tres ejemplos:
- El factor H elimina Bb de la ruta alternativa C3 convertasa rompiendo el bucle de retroalimentación positiva.
- El Factor I inactiva C3b.
- El inhibidor de C1 (C1INH) se une a sitios en C1r y C1s activados, cerrando su actividad proteolítica. Entonces, cuando C1 es activado por complejos antígeno-anticuerpo, solo hay un breve intervalo durante el cual puede escindir C4 y C2 antes de que sea desactivado por C1INH.
Trastornos del Sistema Complemento
Con tantas proteínas involucradas, no es de extrañar que a veces se encuentren en humanos deficiencias heredadas de una u otra. Cuatro ejemplos:
- C3. Una deficiencia hereditaria de C3 predispone a la persona a frecuentes episodios de infecciones bacterianas.
- C2. Curiosamente, los trastornos del complejo inmune, no las infecciones bacterianas, son el principal problema con una deficiencia de C2 (o de uno de los otros componentes “tempranos” como C1q, C1r, C1s o C4). Esto enfatiza el importante papel del sistema del complemento en la eliminación de complejos antígeno-anticuerpo. Una deficiencia de C2 (o uno de los otros componentes tempranos) se encuentra frecuentemente en pacientes con el trastorno autoinmune lupus eritematoso sistémico (LES).
- C9. Otra curiosidad: la mayoría de las personas que no pueden producir C9 no tienen más problemas con las infecciones bacterianas que las que pueden. Estudios de laboratorio sugieren que el complejo C5b•6•7•8 por sí mismo es capaz de lisar bacterias aunque no tan eficientemente como C9. Y, de hecho, una deficiencia de C8 se asocia con un riesgo marcadamente aumentado de meningitis bacteriana.
- C1INH. Una deficiencia de C1INH produce angioedema hereditario (HAE).