
17.3B: Principios de Crecimiento de la Población
- Page ID
- 57065

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Controles independientes de densidad sobre el crecimiento de la población
Los caprichos del entorno físico, por ejemplo la sequía, las congelaciones, los huracanes, las inundaciones y los incendios forestales suelen comprobar el crecimiento de la población. No solo pueden limitar el crecimiento de la población, sino que a menudo conducen a las poblaciones existentes muy por debajo de su nivel anterior. (Y también hacen que sea poco probable que muchos animales sobrevivan el tiempo suficiente para mostrar signos de envejecimiento).
Estos factores se describen como independientes de la densidad porque ejercen su efecto independientemente del tamaño de la población cuando se produjo la catástrofe.
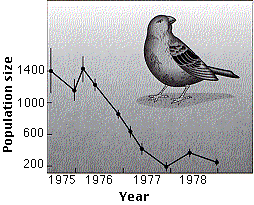

Fig.17.3.2.1 Densidad de población
Esta gráfica (de P. T. Boag y P.R. Grant in Science, 214: 82, 1981) muestra la disminución de la población de uno de los pinzones de Darwin (Geospiza fortis) en Daphne Major, un diminuto (100 acres = 40 hectáreas) miembro de las Islas Galápagos. La disminución (de 1400 a 200 individuos) se produjo debido a una severa sequía que redujo la cantidad de semillas de las que se alimenta esta especie. La sequía terminó en 1978, pero incluso con abundante comida una vez más disponible, la población de pinzones se recuperó poco a poco.
Las caídas catastróficas son particularmente riesgosas para las poblaciones que viven en islas. Cuanto más pequeña es la isla, menor es la población de cada especie en ella, y mayor es el riesgo de que una catástrofe diezme tanto a la población que se extinga. Esto parece ser una razón de la clara relación entre el tamaño de la isla y el número de especies diferentes que contiene.
La gráfica (redibujada de R. H. MacArthur y E. O. Wilson, The Theory of Island Biogeography, Princeton University Press) muestra el número de especies de reptiles y anfibios en diversas islas de las Indias Occidentales. En general, si una isla tiene 10 veces la superficie de otra, contendrá aproximadamente el doble del número de especies.
El mismo principio se aplica a muchos hábitats. En cierto sentido, la mayoría de los hábitats son islas. Una serie de estanques, una cordillera de cimas montañosas, arboledas dispersas de cítricos, incluso árboles individuales dentro de una arboleda, todos están conformados por parches de hábitat separados por barreras a la libre migración de sus habitantes.
Esto tiene importancia tanto práctica como teórica. A medida que crece la población humana, se limpian selvas para la agricultura, se pavimentan granjas para centros comerciales, se embaucan ríos para energía hidroeléctrica y riego, etc. Aunque se están estableciendo santuarios de vida silvestre, deben hacerse lo suficientemente grandes para que puedan soportar poblaciones lo suficientemente grandes como para sobrevivir controles independientes de la densidad cuando golpean.
En 1986, el cierre de una presa en Venezuela inundó más de mil millas cuadradas (>2,500 km 2) convirtiendo cientos de cimas de colinas en islas. Estos variaron en tamaño desde menos de 1 hectárea (2.5 acres) hasta más de 150 hectáreas (370 acres).
Dentro de 8 años:
- Las islas más pequeñas (<1 hectárea) perdieron 75% de las especies que habían vivido allí.
- Cuanto más grande es la isla, menos especies perdió.
- Pero todas las islas -incluso las más grandes- perdieron a sus principales depredadores; es decir, carnívoros como pumas, jaguares (imagen) y águilas en los extremos de las cadenas alimentarias.
- Aquellas especies animales que sí permanecieron -en su mayoría herbívoros y pequeños carnívoros- aumentaron enormemente sus poblaciones debido a
- una reducción en la competencia por los recursos
- ya no ser comido por los depredadores
Comprobaciones del crecimiento poblacional dependientes de la densidad
Competencia Intra Específica
La competencia intraespecífica es competencia entre miembros de la misma especie.
En el verano de 1980, gran parte del sur de Nueva Inglaterra fue golpeada por una infestación de la polilla gitana (Porthetria dispar). A medida que pasaba el verano,
- las larvas (orugas) pupan
- los adultos eclosionados apareados
- las hembras pusieron masas de huevos (cada masa contiene varios cientos de huevos) en prácticamente todos los árboles de la región
- A principios de mayo de 1981, las orugas jóvenes que eclosionaron de estos huevos comenzaron a alimentarse y a mudar.
- Los resultados fueron dramáticos:
- En 72 horas, un árbol de haya de 50 pies o un pino blanco de 25 pies quedaría completamente defoliado.
- Grandes parches de bosque comenzaron a tomar una apariencia invernal con sus esqueletos de ramas desnudas.
- De hecho, la infestación fue tan pesada que muchos árboles fueron completamente defoliados antes de que las orugas pudieran completar su desarrollo larvario.
- El resultado: una muerte masiva de los animales; muy pocos lograron completar la metamorfosis.
Aquí, entonces, fue un ejemplo dramático de cómo la competencia entre miembros de una especie por un recurso finito -en este caso, la comida- provocó una fuerte caída de la población.
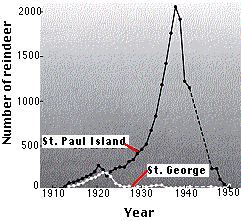
El efecto fue claramente dependiente de la densidad. Las menores densidades poblacionales del verano anterior habían permitido que la mayoría de los animales completaran su ciclo de vida. La gráfica muestra un accidente poblacional similar; en este caso de renos en dos islas del mar de Bering. Se desconoce por qué la población de la Isla San Pablo pasó por un ciclo de auge y caída mucho más severo que el de la Isla San Jorge. Muchas poblaciones de roedores (por ejemplo, lemmings en el Ártico) pasan por tales ciclos de auge y caída.
Competencia Inter específica
Todos los requerimientos ecológicos de una especie constituyen su nicho ecológico. El requerimiento dominante suele ser alimento, pero otros, como los sitios de anidación y un lugar al sol (para las plantas) también pueden ser importantes. Cuando dos especies comparten nichos ecológicos superpuestos, pueden verse forzadas a competir por el (los) recurso (s) de ese nicho. Esta competencia interespecífica es otra comprobación dependiente de la densidad sobre el crecimiento de una o ambas poblaciones.
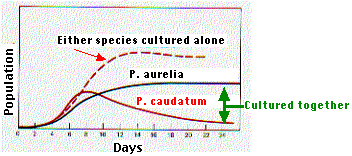
Como tantos factores en ecología, la competencia interespecífica se estudia más fácilmente en el laboratorio que en el campo. Esta gráfica (basada en el trabajo de G. F. Gause) muestra el efecto de la competencia interespecífica sobre el tamaño poblacional de dos especies de paramecia, Paramecium aurelia y Paramecium caudatum.
Cuando cualquiera de las especies fue cultivada sola —con alimentos frescos añadidos regularmente— la población creció exponencialmente al principio y luego se estabilizó.
Sin embargo, cuando las dos especies se cultivaron juntas, P. caudatum demostró ser el competidor más débil. Después de una breve fase de crecimiento exponencial, su población comenzó a declinar y finalmente se extinguió. La población de P. aurelia alcanzó una meseta, pero mientras P. caudatum permaneciera, ésta estuvo por debajo de la densidad poblacional que logró al crecer sola.
El hábitat de la mayoría de las poblaciones naturales es mucho más complejo que un recipiente de cultivo. En un hábitat natural, la especie con ventaja competitiva en una parte del hábitat podría estar en desventaja en otra. Además, la presencia de depredadores y parásitos limitaría el crecimiento poblacional tanto de las especies más exitosas como de las menos exitosas. Entonces, en un entorno natural, el competidor menos efectivo no suele ser impulsado a la extinción.
Desplazamiento de personajes: una respuesta evolutiva que disminuye la competencia interespecífica
Cuando dos especies estrechamente relacionadas se encuentran ocupando la misma ubicación geográfica (simpátrica), sus requerimientos para las necesidades de la vida son probablemente tan similares que se ven forzadas a una intensa competencia interespecífica. Esto puede llevar a uno de los dos posibles resultados.
- La competencia puede ser tan intensa que una especie se elimina por completo; es decir, ahí se ve impulsada a la extinción.
- Alternativamente, el aumento de la presión de selección puede conducir al desplazamiento del carácter, la divergencia evolutiva de un rasgo que ambos utilizan para explotar los recursos de los que ambos dependen. Dicho desplazamiento de personajes disminuye la competencia entre ellos.
En los últimos años, los equipos de investigación liderados por Peter y Rosemary Grant han podido observar el desplazamiento de personajes tal como ocurrió entre dos especies de pinzones de Darwin. Esta es su historia.
- Durante muchos años, G. fortis, el pinzón de tierra mediana, vivió en Daphne Major sin sus parientes, G. fulginosa, el pinzón de tierra pequeño, y G. magnirostris, el grande.
- No obstante, en 1982 llegó G. magnirostris y a lo largo de los años siguientes creció en población.
- Fueron buenos años, con mucha comida, y las dos especies coexistieron muy bien.
- Sin embargo, una severa sequía en 2003-2004, redujo drásticamente los alimentos disponibles, incluidas las semillas grandes por las que compitieron intensamente los magnirostris y los miembros de pico más grande de la población fortis.
- El resultado fue una muerte masiva de ambas especies con los miembros de pico más grande de la población fortis sufriendo mayor mortalidad que los miembros de pico más pequeño de la especie.
- La descendencia de los supervivientes del fortis, nacida en 2005, presentaba picos que promediaban más de 5% menores que el promedio de la población parental. (Después de la sequía de 1977, cuando G. magnirostris no estaba en la isla para competir con fortis, su tamaño de pico aumentó en lugar de disminuir).
- Este estudio sobre el desplazamiento de carácter se describe en la edición del 14 de julio de 2006 de Science.
Competencia Reproductiva
La disminución de las tasas de natalidad también conduce a un crecimiento demográfico
Sabemos que los humanos toman decisiones deliberadas de planificación familiar, pero también se encuentra un comportamiento análogo en otros animales.
- Las moscas de la fruta que viven en condiciones abarrotadas ponen menos huevos
- Las ratas de laboratorio en un área confinada pronto alcanzan un tamaño de población estable a pesar de que hay abundante comida disponible. La causa principal es un fuerte aumento de la mortalidad infantil. La reducción de la atención materna e incluso el canibalismo cobran un alto precio al recién nacido.
- La reina melífera regula su tasa de puesta de huevos a la disponibilidad de alimentos: reduciéndola durante periodos de escasa floración y cesando por completo a finales del verano.
Una alternativa para limitar el número de crías por par de padres es limitar el número de padres. Algunos mamíferos y aves logran esto estableciendo territorios de reproducción. Cada par de apareamiento ocupa un área de un tamaño suficiente para suplir sus necesidades incluyendo las de sus crías. Uno o ambos miembros defienden este territorio contra la intrusión de otros miembros de la misma especie. Este comportamiento no sólo asegura que no se rebasarán los recursos de los que dependen sino que se podrá mantener a la población bajo control impidiendo la reproducción entre sus miembros excedentes.
Las convenciones sociales entre los seres humanos (por ejemplo, actitudes sobre la edad adecuada para contraer matrimonio y el tamaño deseable de la familia) también tienen una marcada influencia en las tasas de natalidad. Sin embargo, las convenciones sociales -y las técnicas anticonceptivas que pueden complementarlas- han tenido más éxito en reducir las tasas de natalidad entre las personas que menos lo necesitan. En los países más pobres, el matrimonio precoz, el deseo de familias numerosas y la falta de empleo confiable de métodos anticonceptivos son comunes.
Migración
La migración suele ser un factor importante dependiente de la densidad para reducir las poblaciones. A medida que aumenta la población, muchos de sus integrantes emigran.
Depredación

A medida que aumenta una población, sus depredadores son capaces de cosecharla con mayor facilidad. Estas gráficas (basadas en datos de Crombie, A. C., Journal of Animal Ecology, 16: 44, 1947) muestran los cambios poblacionales entre los escarabajos harineros cultivados en harina simple (izquierda) y en harina que contiene trozos de tubo de vidrio.
Cada cultivo se inició con cuatro adultos de cada especie. En medio llano, después de un brote inicial de ambas poblaciones, Tribolium continuó expandiendo su número mientras que la población de Oryzaephilus disminuyó y finalmente fue impulsada a la extinción (izquierda). Varios factores estaban en el trabajo, pero la depredación fue, con mucho, la más importante.
- Los adultos de Tribolium se alimentan vorazmente de los huevos y pupas de Oryzaephilus.
- Pero los adultos de Oryzaephilus no se alimentan tan vigorosamente de huevos de Tribolium y no comen sus larvas en absoluto.
Los tubos de vidrio proporcionaron un refugio para algunas larvas de Oryzaephilus, lo que les permitió completar su ciclo de vida. Esta reducción en la intensidad de la depredación permitió que las dos poblaciones convivieran indefinidamente (derecha).
Parasitismo
Los parásitos pueden pasar de hospedador a hospedador más fácilmente a medida que aumenta la densidad poblacional del huésped. Por esta razón, las epidemias entre los humanos son particularmente severas en las ciudades. De hecho, durante la mayor parte del período desde que los humanos comenzaron a vivir en las ciudades, las poblaciones de las ciudades se han mantenido sólo a través de la inmigración continua del campo. No hasta el desarrollo de saneamiento comunitario, inmunización y otras medidas de salud pública, las ciudades evitaron periódicamente caídas bruscas de población como consecuencia de epidemias.
Las epidemias recurrentes de la “muerte negra” en Europa que comenzaron en el siglo XIV provocaron una fuerte disminución de la población. En tan solo 3 años (1348—1350), al menos una cuarta parte de la población de Europa murió a causa de la enfermedad (probablemente peste). Más recientemente, se cree que la gran pandemia de influenza de 1918-1919 mató a más de 20 millones de personas en todo el mundo.
El pinzón común, Carpodacus mexicanus, —originario del oeste de Norteamérica— es un inmigrante reciente al este de Estados Unidos donde es parasitado por un micoplasma que reduce la vida útil y la fecundidad de las aves. Los datos recopilados por observadores aficionados de aves muestran que la llegada de la enfermedad (a mediados de los 90) a zonas con una alta población de aves hizo que su número disminuyera más que en regiones de poblaciones bajas de pinzones. Cualquiera que sea el valor inicial, todas las poblaciones infectadas terminaron con poblaciones similares. Este es un claro ejemplo del efecto dependiente de la densidad del parasitismo en una población.
Ciclos de población
Algunas poblaciones pasan por periodos repetidos y regulares de auge seguidos de busto.
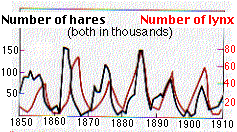
Esta gráfica muestra las fluctuaciones cíclicas a 10 años en las poblaciones (medidas contando las pieles ofrecidas a la venta en los puestos comerciales de la Bahía de Hudson en Canadá) de la liebre variable (“conejo raqueta de nieve”) y su depredador principal, el lince, de 1850 a 1910. El tamaño de la población de lince dependía estrechamente del tamaño de su población presa (liebre). Los factores que provocaron que la población de liebre pasara por sus ciclos de auge y caída aún se debaten, pero la depredación por linces probablemente fue solo un factor.
Estudios de campo recientes han proporcionado respuestas más claras para otras tres poblaciones cíclicas, topillos (un pequeño roedor) en Finlandia, el urogallo rojo en Escocia y lemmings (otro pequeño roedor) en Groenlandia.
Tobillos
La población de ratones en Finlandia pasa regularmente por ciclos de auge y caída de 3 años. Cuando Korpimäki y Norrdahl sacaron a todos sus depredadores (tanto mamíferos como aves) de sus áreas de prueba, los ciclos cesaron. Aquí, entonces, los ciclos fueron impulsados por la verificación de depredación dependiente de la densidad.
Urogallo rojo
La población de urogallo rojo en Escocia pasa por ciclos de 4 a 8 años. De pico a mínimo, la población puede disminuir por un factor de 1000. Estos ciclos no parecen ser causados por la caza de esta popular ave de caza.
Las aves son parasitadas por un nematodo, y las aves infectadas tienen menor fecundidad (tasas de natalidad bajas) y mayor mortalidad (tasas de mortalidad arriba) que las aves no infectadas.
P. J. Hudson y sus colegas trataron a gran número de aves en varias áreas de prueba con un medicamento para prevenir o curar una infección. Las poblaciones en las áreas de tratamiento dejaron de ciclar. No fue necesario tratar a todas las aves; 20% de ellas parecen suficientes para prevenir epidemias (así como la inmunización de humanos no tiene que llegar al 100% para poner fin a la transmisión de patógenos).
Aquí, entonces, los ciclos fueron impulsados por el chequeo de parasitismo dependiente de la densidad.
Lemmings
Un estudio de 15 años de la población de lemmings en el noreste de Groenlandia fue reportado por Gilg, O., et al. , en Ciencia, 31 de octubre de 2003. Estos trabajadores demostraron que la población lemming sube y baja con un ciclo de 4 años. La población de la comadreja de cola corta (también conocida como armiño, armiño), el principal depredador del lemming, también lo hace pero con un retraso de 1 año por detrás de la población lemming.
Debido a este rezago, cabría esperar que la población lemming continuara superando a la población de comadrejas hasta que los lemmings chocaran con la capacidad de carga de su entorno (por ejemplo, disponibilidad de alimentos y sitios de anidación). Pero esto no ocurre porque a medida que crece la población lemming, otros depredadores (por ejemplo, zorros y búhos) cambian su dieta a favor de los lemmings.
A medida que la población lemming comienza entonces a disminuir,
- estos depredadores flexibles regresan a sus antiguas fuentes de alimentos mientras
- cuanto más comadrejas “quisquillosas” disminuyen en número como su única fuente de alimento, los lemmings, tienen.
La capacidad de carga del medio ambiente (K)
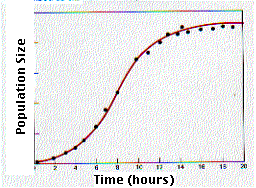
Esta gráfica muestra el crecimiento de una población de levaduras en cultivo. Después de un periodo de crecimiento exponencial, el tamaño de la población comienza a nivelarse y pronto alcanza un valor estable. Este tipo de curva de crecimiento se llama sigmoide o en forma de S. Si agregamos medio de cultivo fresco al recipiente, el crecimiento exponencial se reanuda hasta alcanzar una nueva meseta más alta. Evidentemente, la tasa de crecimiento (r) disminuye a medida que la densidad de la población se acerca a cierto valor limitante.
Cuando r = 0, dN/dt = 0 y la población deja de crecer. Las levaduras han alcanzado un crecimiento poblacional cero o ZPG.
Las causas:
- quedarse sin comida
- acumulación de etanol. (Cuando su concentración alcanza el 12— 14%, la levadura muere (lo que explica el contenido máximo de alcohol de las bebidas alcohólicas naturales como el vino).
El valor limitante de la población que puede ser soportada en un ambiente particular se denomina su capacidad de carga y se designa como K. Cuando la población está muy por debajo de K, su crecimiento es exponencial, pero a medida que la población se acerca a K, comienza a encontrar una "resistencia ambiental” cada vez más fuerte. Usemos la expresión
K − N
K
como un “factor de realización del crecimiento”, es decir, un factor que representa el grado en que la población puede realmente realizar su tasa máxima de incremento posible. Introduciendo este factor en nuestra ecuación de crecimiento original (exponencial), obtenemos
|
dN = rN dt |
( | K − N K |
) |
La ecuación nos dice que
- Si el tamaño de la población (N) está muy por debajo de la capacidad de carga del ambiente (K), el factor de realización del crecimiento será cercano a 1, y la población mostrará un crecimiento exponencial.
- Pero a medida que N comienza a acercarse a K, el factor de realización del crecimiento se acerca a cero, y la tasa de crecimiento de la población cae a cero:
dN = 0 = "ZPG" (crecimiento poblacional cero)
dt
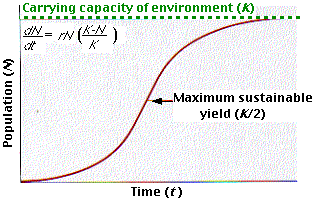
Trazar el crecimiento de una población desde un factor de realización de crecimiento inicial de 1 hasta un factor final de 0 produce una curva como esta, llamada curva de crecimiento logístico o curva de crecimiento en forma de S. Aunque es poco probable que las poblaciones reales sigan exactamente la curva teórica de crecimiento logístico, la curva puede proporcionarnos una valiosa guía en el manejo de las poblaciones.
Ejemplo 1: La curva logística te dice que es poco probable que deshagas tu casa de una gran población de ratas colocando trampas para ratas. No importa cuántas pongas, la r para ratas es tan alta (quizás 0.0147 por día) que se reproducirán más rápido de lo que puedes atraparlas. Lo que debes hacer en cambio es evitar que metan comida dentro y alrededor de tu casa. Con un K muy reducido, su población disminuirá.
Ejemplo 2: Lo contrario del problema de las plagas es cómo evitar que las especies en peligro de extinción se extingan. Pero prohibir la caza no tendrá un impacto apreciable si el hábitat del que depende esa especie para su K —pastos o bosques o lo que sea— desaparece bajo el estacionamiento de una plaza comercial.
Ejemplo 3: Los métodos modernos de pesca intensiva han producido repetidamente descensos ominosos en la captura de muchas especies ya que las poblaciones no han podido mantenerse. La curva logística proporciona un objetivo para el manejo de las pesquerías: cosechar solo a una tasa tal que la población se mantenga en K /2. En este tamaño, la población es capaz de crecer más rápidamente. El valor K /2 se conoce como el rendimiento máximo sustentable.
r -estrategas y K -estrategas
R-estrategas
Una vez aré un campo viejo y permití que estuviera en barbecho. En la primera temporada creció una gran cosecha de ragweed.
La ragweed está bien adaptada para explotar su entorno a toda prisa, antes de que los competidores puedan establecerse. Crece rápidamente y produce una gran cantidad de semillas (después de liberar su polen, la perdición de muchos enfermos de fiebre del heno). Debido a que el enfoque de la ragweed para la supervivencia continua es a través de una reproducción rápida, es decir, un alto valor de r, se le llama r -estratega. Otras malezas, muchos insectos y muchos roedores también son r -estrategas. Si el hecho, si consideramos un organismo una plaga, probablemente sea un r -estratega.
En general, los r -estrategas comparten una serie de características:
- Por lo general, se encuentran en hábitats perturbados y/o transitorios. En la segunda temporada de mi campo, las gramíneas perennes y las flores silvestres habían producido una densa alfombra de vegetación mixta y no se encontraba una planta de ragweed.
- Tienen períodos de vida cortos. El ratón de la casa, con una vida útil máxima de 3 años, es un r -estratega.
- Comienzan a criar temprano en la vida.
- Suelen tener tiempos de generación cortos; es decir, tienen periodos de gestación cortos y pronto están listos para producir otra cosecha de jóvenes. La mosca doméstica puede producir 7 generaciones cada año (cada una de aproximadamente 120 jóvenes).
- Producen un gran número de crías. La ostra americana, liberando un millón de huevos en una temporada, es un r -estratega. La mayoría de sus crías morirán, pero el tamaño de su producción aumenta la probabilidad de que algunas crías se dispersen a nuevos hábitats.
- Ellos cuidan poco a sus crías, y la mortalidad infantil es enorme. Si trazamos una curva de supervivencia para un estratega r, es apto para tomar la forma de la curva etiquetada D. Aunque los humanos no son estrategas r, la mayor tasa de natalidad en algunos países bien puede ser una respuesta a sus mayores tasas de mortalidad infantil (curva B).
- Cuentan con medios eficientes de dispersión a nuevos hábitats.
Para los r -estrategas, los alelos que potencien cualquiera de los rasgos enumerados anteriormente se verán favorecidos por la selección natural. De ahí que se dice que los r -estrategas son producto de r -selección.
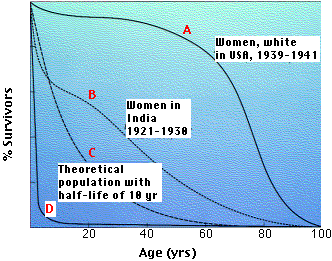
La gráfica muestra 4 curvas representativas de supervivencia. El eje vertical da la fracción de supervivientes a cada edad.
- La curva A es característica de organismos que tienen baja mortalidad hasta finales de la vida cuando el envejecimiento pasa factura.
- La cura B es típica de poblaciones en las que factores como el hambre y la enfermedad oscurecen los efectos del envejecimiento, y la mortalidad infantil es alta.
- La curva C es una curva teórica para organismos para los que la probabilidad de muerte es igual en todas las edades. Este podría ser el caso de organismos que no envejecen (algunos peces) o aquellos que sufren una mortalidad aleatoria severa a lo largo de la vida (por ejemplo, muchos pájaros cantores). Los estrategas K suelen tener curvas de supervivencia en algún lugar entre A y C.
- La curva D es típica de organismos, ostras por ejemplo, que producen un gran número de crías acompañadas de altas tasas de mortalidad infantil. Muchos r -estrategas tienen tal curva.
K-estrategas
Cuando un hábitat se llena de una colección diversa de criaturas que compiten entre sí por las necesidades de la vida, la ventaja cambia a K -estrategas. Los estrategas K tienen poblaciones estables cercanas a K. No hay nada que ganar de una r alta. La especie se beneficiará más por una adaptación cercana a las condiciones de su entorno.
Por lo general, los estrategas K comparten estas cualidades:
- Por lo general, se encuentran en hábitats estables. La mayoría de las especies en un bosque maduro serán K -estrategas.
- Tienen una larga vida útil. El elefante y la tortuga son K -estrategas.
- Comienzan a reproducirse más tarde en la vida.
- Suelen tener tiempos de generación largos. Se necesitan 9 meses para producir un bebé humano.
- La mayoría produce pequeñas cantidades de descendencia. Las aves son estrategas K, la mayoría de las especies producen menos de una docena de crías cada año.
- Ellos cuidan bien a sus crías. La mortalidad infantil tiende a ser baja. Si trazamos una curva de supervivencia para un estratega K, generalmente se encuentra en algún lugar entre la curva A (arriba), donde la mayoría de la población muere en la vejez, y la curva C, donde todas las edades corren el mismo riesgo de ser derribadas por peligros aleatorios.
- Los estrategas K generalmente han evolucionado de tal manera que se vuelven cada vez más eficientes para explotar una porción cada vez más estrecha de su entorno. Por lo tanto, no es de extrañar que muchas especies en peligro de extinción sean K -estrategas.
Para los K-estrategas, los alelos que potencian su capacidad para explotar los recursos de su hábitat; es decir, para aumentar la capacidad de carga, K, de su entorno, se verán favorecidos por la selección natural. De ahí que se dice que los K -estrategas son producto de K -selección.
La densidad poblacional puede provocar cambios en la estrategia
Un equipo del campus Santa Cruz de la Universidad de California (Sinervo et al., en la edición del 21 de agosto de 2001 de Nature) estudió los ciclos de auge y busto del lagarto nativo de manchas laterales. Encontraron que la población de lagartos pasó por ciclos de dos años de auge y busto.
Año 1 = Boom
- Una población baja de adultos
- viviendo muy por debajo de la capacidad de carga (K) de su entorno
- produjo un gran número de jóvenes (una estrategia r)
- lo que lleva a un rápido hacinamiento y
Año 2 = Busto
- Una gran población de adultos
- viviendo cerca o por encima de la K de su entorno
- produjo menos jóvenes sobrevivientes
- lo que lleva a una fuerte disminución de la población y
Año 3 = otro año Boom, y así sucesivamente.
También encontraron que la población es polimórfica que contiene:
- hembras con gargantas anaranjadas que producían hasta 5 nidadas de huevos (promediando 6 huevos por nidada) por temporada. Se necesitan muchas reservas de alimentos para hacer huevos y los huevos de estas prolíficas hembras de garganta naranja tendieron a ser más pequeños —y a eclosionar en lagartijas más pequeñas— que los de la
- hembras con gargantas amarillas. Estas hembras tienden a poner menos huevos, pero más grandes, y las lagartijas jóvenes que eclosionan de ellas son más grandes que las producidas por las madres de garganta naranja.
Como predijeron, resultó que:
- Los lagartos de garganta naranja son r -estrategas. En los años de auge, tuvieron más éxito que el amarillo. La explosión poblacional de lagartijas jóvenes producidas por ellos provocó el busto del próximo año.
- Los lagartos de garganta amarilla son K -estrategas. Al producir un número menor de lagartos más grandes, tuvieron más éxito en dejar crías sobrevivientes para sentar las bases para el próximo año de auge.
Aquí, entonces, la competencia intraespecífica ha creado un ciclo poblacional favoreciendo alternativamente a los r -estrategas y a los K -estrategas.