
18.1: Evolución y Adaptación
- Page ID
- 57291

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
En 1859 el naturalista inglés Charles Darwin publicó El origen de las especies. El libro contenía dos grandes argumentos: Primero, Darwin presentó una gran cantidad de evidencia de la evolución. Dijo que todos los seres vivos en la tierra hoy en día son los descendientes -con modificaciones- de especies anteriores. En segundo lugar, propuso un mecanismo -selección natural- para explicar cómo se da la evolución.
La evolución involucra dos fenómenos interrelacionados: la adaptación y la especiación. En la adaptación, a lo largo del tiempo, las especies modifican sus fenotipos de manera que les permiten tener éxito en su entorno. En especiación, a lo largo del tiempo, el número de especies se multiplica; es decir, una sola especie puede dar lugar a dos o más especies descendientes. De hecho, Darwin sostuvo que todas las especies están relacionadas; es decir, dos especies cualesquiera en la tierra hoy en día han compartido un ancestro común en algún momento de su historia.
Selección Natural
- Los seres vivos producen más descendencia de lo que pueden soportar los recursos finitos que tienen a su disposición.
- Así los seres vivos enfrentan una lucha constante por la existencia.
- Los individuos en una población varían en sus fenotipos.
- Parte de esta variación es heredable; es decir, es un reflejo de variaciones en el genotipo.
- Aquellas variantes mejor adaptadas a las condiciones de su vida tienen más probabilidades de sobrevivir y reproducirse (“supervivencia del más apto”).
- En la medida en que sus adaptaciones sean heredables, serán transmitidas a su descendencia.
Las fuerzas de selección natural actúan sobre los fenotipos, pero sólo si hay un cambio en los genotipos de una población se ha producido evolución.
La medida de la “aptitud”
La aptitud física es una medida del éxito reproductivo. Aquellos individuos que dejan el mayor número de crías maduras son los más aptos. Esto se puede lograr de varias maneras:
- Supervivencia (selección de mortalidad)
- Éxito en el apareamiento (selección sexual)
- Tamaño de la familia (selección de fecundidad)
Supervivencia
Cualquier rasgo que promueva la supervivencia -al menos hasta que terminen los años reproductivos- aumenta la condición física. Tales rasgos son adaptaciones.
Selección Sexual
En la selección sexual, un sexo -generalmente el femenino- elige entre los machos disponibles. Cualquier rasgo heredado que mejore el éxito de apareamiento de ciertos individuos se volverá más pronunciado en las generaciones venideras. Algunos ejemplos:
- Cuando están listas para aparearse, las hembras de tres espinas espinosas (peces) eligen machos con muchos alelos MHC Clase II sobre los machos con menos alelos. Los alelos de clase II codifican las proteínas que presentan antígenos al sistema inmune. Presumiblemente, cuanto más tengas, mayor será la diversidad de antígenos parasitarios que tu sistema inmunitario pueda reconocer y defender.Las hembras distinguen entre los machos por moléculas solubles (“olores”) que los machos liberan al agua. Se desconoce cómo estos “olores” son controlados por los alelos del MHC.
- Una cultura de Drosophila establecida con igual número de moscas de ojos rojos y de ojos blancos de ambos sexos acabará, después de 25 generaciones más o menos, teniendo solo moscas de ojos rojos (las “normales”) en ella. Esto a pesar de que las moscas de ojos blancos son igual de saludables y viven tanto como las moscas de ojos rojos, es decir, son iguales en términos de supervivencia. Pero, como resulta, no sólo las hembras de ojos rojos prefieren a los machos de ojos rojos, sino que las hembras de ojos blancos también.
En otros casos de selección sexual, un fenotipo prefiere aparearse con otros del mismo fenotipo. A esto se le llama apareamiento selectivo.
Selección de fecundidad
La producción de un gran número de crías maduras es una medida de aptitud. Yo enfatizo madurar porque solo ellos pueden transmitir estos rasgos a otra generación. Algunas formas de hacer esto:
- Crianza más temprana. Si algunas hembras maduran sexualmente antes que otras, aumentan sus posibilidades de dejar descendencia.
- Para algunas especies (por ejemplo, peces, ostras), que proporcionan poco o ningún cuidado a sus crías, la aptitud se mide por el número de huevos fertilizados que producen.
- Para las especies (como nosotros) que cuidan a sus crías, la selección actúa para reducir el tamaño de la familia (hasta cierto punto). Un amplio estudio realizado en Utah (Estados Unidos) demostró que en el siglo XIX, las familias con menos hijos tenían más nietos sobrevivientes.
Todas las fuerzas de selección natural señaladas anteriormente trabajan en individuos. Pero cada vez hay más evidencias de que la selección natural también puede actuar sobre los grupos. La selección natural que parece funcionar en contra del beneficio de algunos individuos al tiempo que mejora las perspectivas de sus familiares se llama selección de parientes. Se discute en una página separada.
¿Están los humanos exentos de la selección natural?
Se ha argumentado que los avances en medicina, saneamiento, etc. han alejado a los humanos de los rigores de la selección natural. Probablemente hay algo de verdad en esto, pero considera que de todos los óvulos humanos que se fertilizan, menos de la mitad jamás se reproducirán. Los demás se eliminan de la siguiente manera:
- Selección de mortalidad
- Aproximadamente 30% de los embarazos terminan por aborto espontáneo de embriones y fetos o por muerte fetal.
- La muerte en la infancia y la infancia reclama otro 5% o más.
- Selección sexual
- Otro 20% sobrevivirá hasta la edad adulta pero nunca se casará.
- Selección de fecundidad
- De los que sí se casan, el 10% no tendrá hijos.
Variación continua
La mayoría de los rasgos en una población como la estatura y el peso corporal varían de manera continua de un extremo a otro.
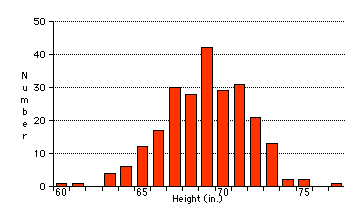
Una gráfica de la distribución del rasgo en una población a menudo produce una curva en forma de campana como esta que muestra la distribución de las alturas entre un grupo de estudiantes de último año de secundaria varones. Tal distribución podría surgir de factores ambientales -quizás la variación continua de altura en los niños es simplemente el resultado de la variación en su dieta a medida que crecían o factores genéticos- los padres altos tienden a tener hijos altos o -muy probablemente- ambos.
Heredabilidad
Se puede ordenar la contribución relativa de los factores genéticos y ambientales comparando el rango de un rasgo en la descendencia en comparación con el valor promedio de ese rasgo en sus padres. Si la descendencia de padres seleccionados ocupa el mismo rango que toda la población, los factores ambientales están trabajando solos. El rasgo tiene una heredabilidad cero. Ejemplo: La longitud de las semillas de una cepa pura de frijol puede variar en varios milímetros. Sin embargo, si se aparean frijoles extra grandes, el nuevo cultivo no muestra ningún cambio a un tamaño mayor. Entonces la heredabilidad de la longitud es cero. Por otro lado, si las crías de dos ratones extragrandes son tan grandes como lo son, los genes probablemente estén trabajando. Se dice que el rasgo tiene una heredabilidad de 1.

Los efectos de la selección en las poblaciones
Las presiones de selección natural pueden afectar la distribución de fenotipos en una población de varias maneras.
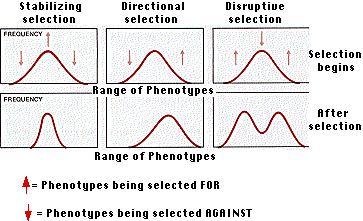
Selección estabilizadora
La selección natural a menudo trabaja para eliminar a los individuos en ambos extremos de una variedad de fenotipos, lo que resulta en el éxito reproductivo de aquellos que están cerca de la media. En tales casos, el resultado es mantener el status quo. No siempre es fácil ver por qué ambos extremos deben ser discapacitados; quizás la selección sexual o la responsabilidad ante la depredación está en el trabajo. En cualquier caso, la selección estabilizadora es común. En humanos, por ejemplo, la incidencia de mortalidad infantil es mayor tanto para bebés muy pesados como para bebés muy ligeros.
Selección Direccional
Una población puede encontrarse en circunstancias en las que los individuos que ocupan un extremo en el rango de fenotipos son favorecidos sobre los demás. Desde 1973, Peter y Rosemary Grant -ayudados por una sucesión de colegas- han estudiado los pinzones de Darwin en las Islas Galápagos. Cuando las lluvias, y por lo tanto los alimentos, son abundantes, los pinzones molidos tienden a tener una dieta variada, por ejemplo, comen semillas de una variedad de tamaños y muestran una variación considerable en el cuerpo y el tamaño del pico (los picos grandes son mejores para las semillas grandes pero pueden manejar semillas pequeñas así como pequeños picos).

De 1976 a 1977, una severa sequía azotó las islas, prácticamente sin precipitaciones durante más de un año. Esto provocó una disminución precipitada en la producción de las semillas que son el pilar dietético de Geospiza fortis, el pinzón medio molido. La gráfica (de P. T. Boag y P. R. Grant in Science 214:82, 1981) muestra cómo la población disminuyó de 1400 a 200 en la isla de Daphne Major, una minúscula (10 acres = 4 hectáreas) miembro de las Islas Galápagos.
Una de las plantas para superar la sequía produce semillas en frutos grandes y duros que son prácticamente imposibles de comer para las aves con pico menor a 10.5 mm. El muestreo de las aves que murieron así como las que sobrevivieron mostró que las aves más grandes fueron favorecidas sobre las más pequeñas y las que tenían picos más grandes se favorecieron sobre las de menor tamaño.
| Largo del pico (mm) | Profundidad del pico (mm) | |
|---|---|---|
| Aves muertas | 10.68 | 9.42 |
| Sobrevivientes | 11.07 | 9.96 |
Aquí, entonces fue la selección natural en el trabajo. Pero, ¿produjo evolución? La respuesta resultó ser sí. A medida que la población de G. fortis se recuperó después de que regresaran las lluvias, el tamaño corporal promedio y la profundidad del pico de sus crías fueron mayores que antes (un aumento de 4— 5% para la profundidad del pico). La curva acampanada había sido desplazada hacia la derecha — selección direccional.
Más recientemente, los Grants y colegas de la Escuela de Medicina de Harvard han demostrado que
- El ancho y profundidad del pico en los pinzones del suelo se correlacionan con el tiempo y la intensidad de expresión del gen, Bmp4, (que codifica la proteína morfogenética ósea-4) en el tejido que formará el pico superior. La expresión de Bmp4 aparece más temprano en el desarrollo y con mayor intensidad en la Geospiza magnirostris de pico grande (el pinzón de tierra grande) que en sus parientes de pico más pequeño, Geospiza fortis (el pinzón de tierra mediana) y Geospiza fuliginosa (la pequeña pinzón molido). Ver Abzhanov, A., et al., Science, 3 de septiembre de 2004.
- Sin embargo, la longitud del pico se correlaciona con la intensidad de expresión del gen CaM que codifica la calmodulina de la proteína de unión a Ca 2+ en el tejido que formará el pico superior. La expresión de CaM es mucho mayor en el tejido embrionario de los pinzones cactus (G. scandens y G. conirostris, ambos con picos largos) que en sus parientes de pico corto, los pinzones terrestres G. fortis y G. magnirostris. (Véase Abzhanov, A., et al., Nature, 3 de agosto de 2006.)
Melanismo industrial
Muchas especies de polillas en las Islas Británicas comenzaron a tornarse de color más oscuro en el siglo XIX. El ejemplo mejor estudiado es la polilla salpimentada, Biston betularia. La polilla recibe su nombre de las manchas oscuras dispersas en sus alas y cuerpo. En 1849, un mutante negro carbón fue encontrado cerca de Manchester, Inglaterra. Dentro de un siglo, esta forma negra había aumentado al 90% de la población en esta región. La polilla vuela de noche y descansa de día sobre los troncos de los árboles. En zonas alejadas de la actividad industrial, los troncos de los árboles están incrustados de líquenes. Como muestran las fotos, la forma de luz (rodeada en rojo) es prácticamente invisible sobre este fondo.

En zonas donde la contaminación del aire es severa, la combinación de gases tóxicos y hollín ha matado los líquenes y ennegrecido los troncos. Contra tal fondo, la forma de luz destaca bruscamente. La polilla es presa de aves que la arrancan de su lugar de descanso durante el día. En bosques contaminados, la forma oscura tiene muchas más posibilidades de sobrevivir sin ser detectada. Cuando el genetista inglés H. B. D. Kettlewell (quien suministró las fotos) soltó polillas de ambos tipos en el bosque, observó que las aves, efectivamente, comían una fracción mucho mayor de las polillas ligeras que soltaba que de la oscuridad. Desde que se pusieron en marcha programas de reducción de la contaminación después de la Segunda Guerra Mundial, la forma de luz ha estado regresando en las zonas de Liverpool y Manchester.
Selección disruptiva
En algunas circunstancias, los individuos en ambos extremos de una variedad de fenotipos son favorecidos sobre los del medio. Esto se llama selección disruptiva.
Un ejemplo:
Los residuos (“relaves”) de las minas a menudo contienen concentraciones tan altas de metales tóxicos (por ejemplo, cobre, plomo) que la mayoría de las plantas no pueden crecer en ellos. Sin embargo, algunas especies resistentes (por ejemplo, ciertas gramíneas) son capaces de extenderse desde el suelo circundante no contaminado a dichos montones de desechos. Estas plantas desarrollan resistencia a los metales tóxicos mientras que su capacidad de crecer en suelos no contaminados disminuye. Debido a que los pastos son polinizados por el viento, continúa la reproducción entre las poblaciones resistentes y no resistentes. Pero evidentemente, la selección disruptiva está en juego. Las tasas de mortalidad más altas tanto de plantas menos resistentes que crecen en suelo contaminado como de plantas más resistentes que crecen en suelo no contaminado, conducen a una creciente divergencia de las poblaciones en dos subpoblaciones con las manifestaciones extremas de este rasgo.
La importancia evolutiva de la selección disruptiva radica en la posibilidad de que el acervo genético pueda dividirse en dos reservas genéticas distintas. Esta puede ser una forma en la que se formen nuevas especies. La formación de una o más especies a partir de una sola especie precursora se denomina especiación.